Refine search
Actions for selected content:
16123 results in Animal behaviour
1 - Asking Questions about Primates
-
- Book:
- Studying Primates
- Published online:
- 19 September 2019
- Print publication:
- 26 September 2019, pp 1-16
-
- Chapter
- Export citation
Index
-
- Book:
- Studying Primates
- Published online:
- 19 September 2019
- Print publication:
- 26 September 2019, pp 333-342
-
- Chapter
- Export citation
25 - Conclusions
-
- Book:
- Studying Primates
- Published online:
- 19 September 2019
- Print publication:
- 26 September 2019, pp 327-332
-
- Chapter
- Export citation
10 - Finding Out What We Know
-
- Book:
- Studying Primates
- Published online:
- 19 September 2019
- Print publication:
- 26 September 2019, pp 137-146
-
- Chapter
- Export citation
4 - Inclusive Science
-
- Book:
- Studying Primates
- Published online:
- 19 September 2019
- Print publication:
- 26 September 2019, pp 45-52
-
- Chapter
- Export citation
16 - Sampling and Statistical Power
-
- Book:
- Studying Primates
- Published online:
- 19 September 2019
- Print publication:
- 26 September 2019, pp 207-214
-
- Chapter
- Export citation
19 - Collecting Data
-
- Book:
- Studying Primates
- Published online:
- 19 September 2019
- Print publication:
- 26 September 2019, pp 241-246
-
- Chapter
- Export citation
Acknowledgements
-
- Book:
- Studying Primates
- Published online:
- 19 September 2019
- Print publication:
- 26 September 2019, pp xvii-xx
-
- Chapter
- Export citation
17 - Checking Feasibility and Finalising Your Plans
-
- Book:
- Studying Primates
- Published online:
- 19 September 2019
- Print publication:
- 26 September 2019, pp 215-222
-
- Chapter
- Export citation
11 - Reading Journal Articles
-
- Book:
- Studying Primates
- Published online:
- 19 September 2019
- Print publication:
- 26 September 2019, pp 147-156
-
- Chapter
- Export citation
12 - Formulating Hypotheses and Predictions and Designing a Study
-
- Book:
- Studying Primates
- Published online:
- 19 September 2019
- Print publication:
- 26 September 2019, pp 157-166
-
- Chapter
- Export citation
Copyright page
-
- Book:
- Studying Primates
- Published online:
- 19 September 2019
- Print publication:
- 26 September 2019, pp iv-iv
-
- Chapter
- Export citation

Studying Primates
- How to Design, Conduct and Report Primatological Research
-
- Published online:
- 19 September 2019
- Print publication:
- 26 September 2019
Molecular confirmation and description of the larval morphology of Thalassocaris lucida (Dana, 1852) (Decapoda, Caridea, Pandaloidea)
-
- Journal:
- Journal of the Marine Biological Association of the United Kingdom / Volume 99 / Issue 8 / December 2019
- Published online by Cambridge University Press:
- 19 September 2019, pp. 1797-1805
-
- Article
- Export citation
Influence of artisanal fisheries on the behaviour and social structure of Tursiops truncatus in the South-western Gulf of Mexico
-
- Journal:
- Journal of the Marine Biological Association of the United Kingdom / Volume 99 / Issue 8 / December 2019
- Published online by Cambridge University Press:
- 10 September 2019, pp. 1841-1849
-
- Article
- Export citation
-
Behavioural plasticity in animals is tested whenever competitive interactions for space and/or food resources occur between wildlife and human activities. This study uses the concepts of operational and non-operational interactions between bottlenose dolphins (Tursiops truncatus) and artisanal fisheries in Alvarado, to search for differences in behaviour, age structure and group size. We conducted 20 surveys between 2015 and 2016, and recorded 64 groups by means of scan sampling from either a research boat or a fixed vantage point. Average dolphin group size was small (
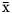 ${\bar{\rm x}}$= 3.2, SD = 2.2 individuals) and fewer individuals were commonly present when interaction with fisheries occurred. Operational interactions were defined within the first 30 m and occurred mainly with lone individuals (54% recorded from the lighthouse and 82% during surveys); this benchmark also accounted for higher frequencies in locomotion and feeding (χ2 = 83.10; df = 7; P < 0.001). We found a higher rate of new behavioural events for dolphin groups furthest from human activities, as well as a decrease in behaviours that imply greater body exposure as dolphins approach the fishing spots. Age structure and dolphin group size were not different during and in the absence of interaction with fisheries, but most interactions involved male dolphins. Behavioural variations in the dolphins' repertoire are likely a strategy to reduce the risk of injuries or death when interacting with human activities; these dolphins seem to have habituated to or at least tolerate fishing activities within the study area, possibly constituting a sex-biased pressure.
${\bar{\rm x}}$= 3.2, SD = 2.2 individuals) and fewer individuals were commonly present when interaction with fisheries occurred. Operational interactions were defined within the first 30 m and occurred mainly with lone individuals (54% recorded from the lighthouse and 82% during surveys); this benchmark also accounted for higher frequencies in locomotion and feeding (χ2 = 83.10; df = 7; P < 0.001). We found a higher rate of new behavioural events for dolphin groups furthest from human activities, as well as a decrease in behaviours that imply greater body exposure as dolphins approach the fishing spots. Age structure and dolphin group size were not different during and in the absence of interaction with fisheries, but most interactions involved male dolphins. Behavioural variations in the dolphins' repertoire are likely a strategy to reduce the risk of injuries or death when interacting with human activities; these dolphins seem to have habituated to or at least tolerate fishing activities within the study area, possibly constituting a sex-biased pressure.
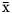
Barnacle cover modifies foraging behaviour of the intertidal limpet Patella vulgata
-
- Journal:
- Journal of the Marine Biological Association of the United Kingdom / Volume 99 / Issue 8 / December 2019
- Published online by Cambridge University Press:
- 10 September 2019, pp. 1779-1786
-
- Article
-
- You have access
- HTML
- Export citation
Monitoring green sea turtles in Brazilian feeding areas: relating body condition index to fibropapillomatosis prevalence
-
- Journal:
- Journal of the Marine Biological Association of the United Kingdom / Volume 99 / Issue 8 / December 2019
- Published online by Cambridge University Press:
- 10 September 2019, pp. 1879-1887
-
- Article
- Export citation
First report of brooding of eggs in the deep-sea genus Anguillosyllis (Annelida: Syllidae)
-
- Journal:
- Journal of the Marine Biological Association of the United Kingdom / Volume 99 / Issue 8 / December 2019
- Published online by Cambridge University Press:
- 10 September 2019, pp. 1775-1777
-
- Article
- Export citation
Estimation of natural mortality in two demersal squat lobster species off Chile
-
- Journal:
- Journal of the Marine Biological Association of the United Kingdom / Volume 99 / Issue 7 / November 2019
- Published online by Cambridge University Press:
- 09 September 2019, pp. 1639-1650
-
- Article
- Export citation
Verification of ageing methods for the burrowing shrimp, Neotrypaea californiensis, using extractable lipofuscin and gastric mill cuticular features
-
- Journal:
- Journal of the Marine Biological Association of the United Kingdom / Volume 99 / Issue 7 / November 2019
- Published online by Cambridge University Press:
- 09 September 2019, pp. 1627-1638
-
- Article
- Export citation
