Refine search
Actions for selected content:
86 results
Shared and distinct alterations of thalamic subregional functional connectivity in early- and late-onset obsessive-compulsive disorder
-
- Journal:
- Psychological Medicine / Volume 55 / 2025
- Published online by Cambridge University Press:
- 01 September 2025, e258
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Progressive gray matter reduction in schizophrenia patients with persistent auditory hallucinations by causal structural covariance network analysis
-
- Journal:
- Psychological Medicine / Volume 55 / 2025
- Published online by Cambridge University Press:
- 29 August 2025, e255
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Exploring connectivity and volume alterations in the Pulvinar’s subnuclei: insights into the neuropathological role in obsessive-compulsive disorder (OCD)
-
- Journal:
- Psychological Medicine / Volume 55 / 2025
- Published online by Cambridge University Press:
- 21 July 2025, e206
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Thalamus as a neural marker of cognitive reserve in bilinguals with frontotemporal dementia
-
- Journal:
- Bilingualism: Language and Cognition , First View
- Published online by Cambridge University Press:
- 22 April 2025, pp. 1-10
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Chapter 40 - The Pathophysiology of Tremor
- from Section 3: - Hyperkinetic Movement Disorders
-
-
- Book:
- International Compendium of Movement Disorders
- Published online:
- 07 January 2025
- Print publication:
- 06 February 2025, pp 499-512
-
- Chapter
- Export citation
Longitudinal association between depressive symptoms and cognitive function: the neurological mechanism of psychological and physical disturbances on memory
-
- Journal:
- Psychological Medicine / Volume 54 / Issue 13 / October 2024
- Published online by Cambridge University Press:
- 14 October 2024, pp. 3602-3611
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Common neural deficits across reward functions in major depression: a meta-analysis of fMRI studies
-
- Journal:
- Psychological Medicine / Volume 54 / Issue 11 / August 2024
- Published online by Cambridge University Press:
- 23 May 2024, pp. 2794-2806
-
- Article
- Export citation
98 Cognitive Outcomes Following Bilateral Focused Ultrasound Thalamotomy for Tremor
-
- Journal:
- Journal of the International Neuropsychological Society / Volume 29 / Issue s1 / November 2023
- Published online by Cambridge University Press:
- 21 December 2023, pp. 197-198
-
- Article
-
- You have access
- Export citation
Mindfulness training and exercise differentially impact fear extinction neurocircuitry
-
- Journal:
- Psychological Medicine / Volume 54 / Issue 4 / March 2024
- Published online by Cambridge University Press:
- 01 September 2023, pp. 835-846
-
- Article
- Export citation
Chapter 14 - Motor Thalamic Interactions with the Brainstem and Basal Ganglia
- from Section 6: - Motor Control
-
-
- Book:
- The Thalamus
- Published online:
- 12 August 2022
- Print publication:
- 01 September 2022, pp 269-283
-
- Chapter
- Export citation
Chapter 16 - The Thalamus in Cognitive Control
- from Section 7: - Cognition
-
-
- Book:
- The Thalamus
- Published online:
- 12 August 2022
- Print publication:
- 01 September 2022, pp 307-323
-
- Chapter
- Export citation
Chapter 10 - Corticothalamic Feedback in Vision
- from Section 5: - Sensory Processing
-
-
- Book:
- The Thalamus
- Published online:
- 12 August 2022
- Print publication:
- 01 September 2022, pp 206-213
-
- Chapter
- Export citation
The thalamus and its subregions – a gateway to obsessive-compulsive disorder
-
- Journal:
- European Psychiatry / Volume 65 / Issue S1 / June 2022
- Published online by Cambridge University Press:
- 01 September 2022, pp. S77-S78
-
- Article
-
- You have access
- Open access
- Export citation
-
Introduction
Higher thalamic volume has been found in children with obsessive-compulsive disorder (OCD) and children with clinical-level symptoms within the general population (Boedhoe et al. 2017, Weeland et al. 2021a). Functionally distinct thalamic nuclei are an integral part of OCD-relevant brain circuitry.
ObjectivesWe aimed to study the thalamic nuclei volume in relation to subclinical and clinical OCD across different age ranges. Understanding the role of thalamic nuclei and their associated circuits in pediatric OCD could lead towards treatment strategies specifically targeting these circuits.
MethodsWe studied the relationship between thalamic nuclei and obsessive-compulsive symptoms (OCS) in a large sample of school-aged children from the Generation R Study (N = 2500) (Weeland et al. 2021b). Using the data from the ENIGMA-OCD working group we conducted mega-analyses to study thalamic subregional volume in OCD across the lifespan in 2,649 OCD patients and 2,774 healthy controls across 29 sites (Weeland et al. 2021c). Thalamic nuclei were grouped into five subregions: anterior, ventral, intralaminar/medial, lateral and pulvinar (Figure 1).
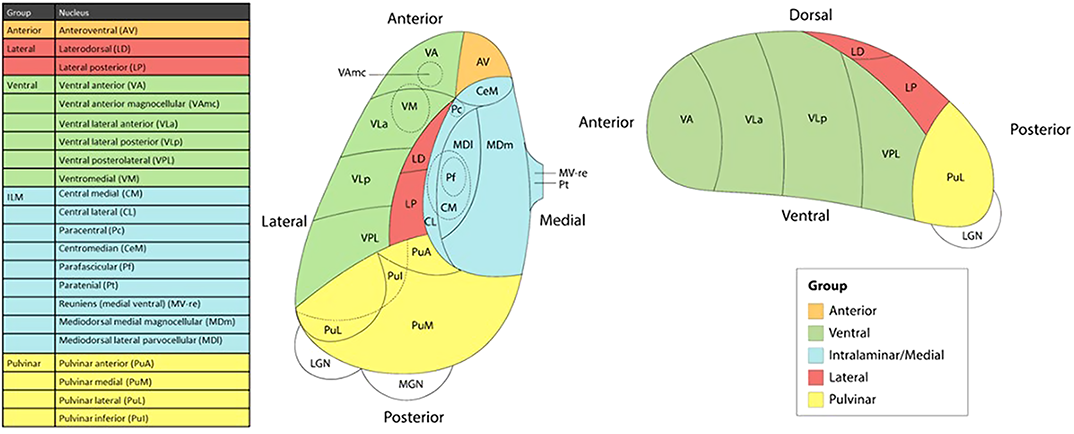 Results
ResultsBoth children with subclinical and clinical OCD compared with controls show increased volume across multiple thalamic subregions. Adult OCD patients have decreased volume across all subregions (Figure 2), which was mostly driven by medicated and adult-onset patients.
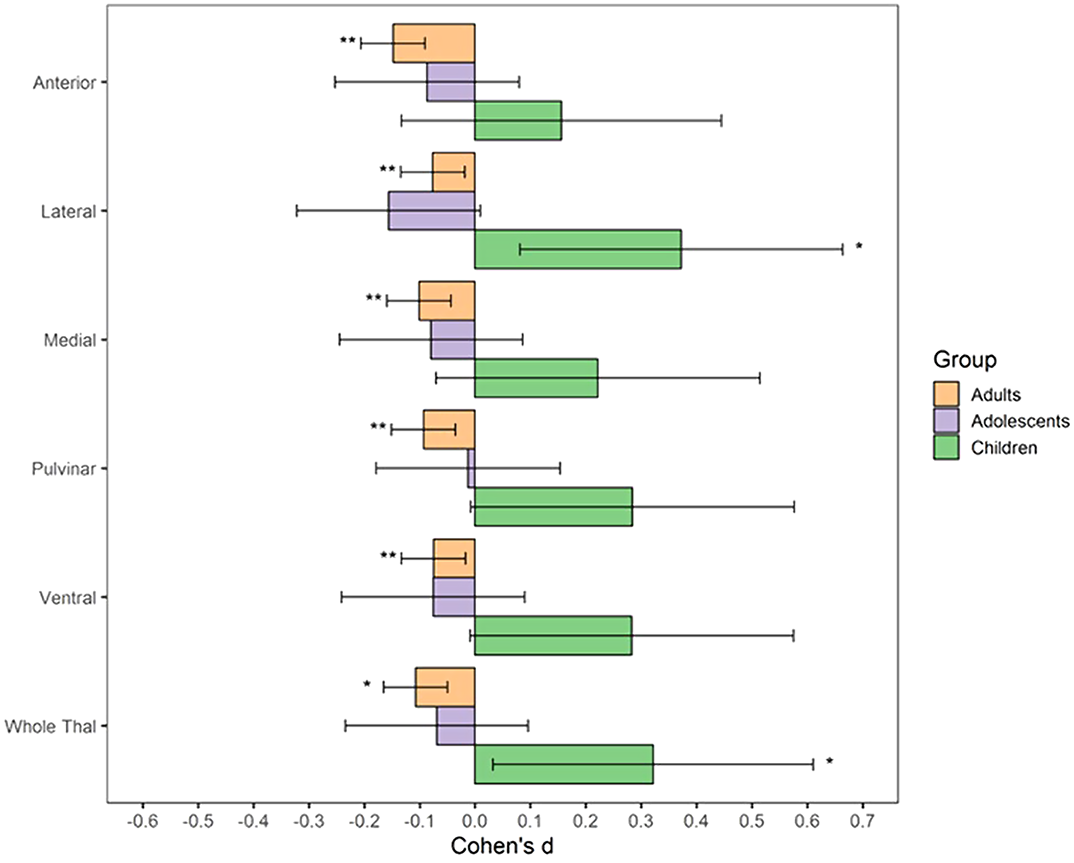 Conclusions
ConclusionsOur results suggests that OCD-related thalamic volume differences are global and not driven by particular subregions and that the direction of effects are driven by both age and medication status.
DisclosureNo significant relationships.


Chapter 9 - Thalamocortical Interactions in the Primary Visual Cortex
- from Section 5: - Sensory Processing
-
-
- Book:
- The Thalamus
- Published online:
- 12 August 2022
- Print publication:
- 01 September 2022, pp 187-205
-
- Chapter
- Export citation
The Clinical and Cognitive Spectrum of Artery of Percheron Infarction: 1-Year Follow-Up
-
- Journal:
- Canadian Journal of Neurological Sciences / Volume 49 / Issue 6 / November 2022
- Published online by Cambridge University Press:
- 10 September 2021, pp. 774-780
-
- Article
-
- You have access
- HTML
- Export citation
Alpha-Mannosidosis: A Novel Cause of Bilateral Thalami and Dentate Nuclei Hyperintensity
-
- Journal:
- Canadian Journal of Neurological Sciences / Volume 49 / Issue 5 / September 2022
- Published online by Cambridge University Press:
- 06 September 2021, pp. 704-705
-
- Article
-
- You have access
- HTML
- Export citation
11 - Processes of Utterance Interpretation: For a Neuropragmatics
- from Part IV - The Processing of Speech Meaning
-
- Book:
- The Study of Speech Processes
- Published online:
- 14 January 2021
- Print publication:
- 21 January 2021, pp 220-238
-
- Chapter
- Export citation
The impact of childhood trauma on thalamic functional connectivity in patients with obsessive–compulsive disorder
-
- Journal:
- Psychological Medicine / Volume 52 / Issue 13 / October 2022
- Published online by Cambridge University Press:
- 20 November 2020, pp. 2471-2480
-
- Article
- Export citation
Stimulation of the Anterior Nucleus of the Thalamus for Epilepsy: A Canadian Experience
- Part of
-
- Journal:
- Canadian Journal of Neurological Sciences / Volume 48 / Issue 4 / July 2021
- Published online by Cambridge University Press:
- 16 October 2020, pp. 469-478
-
- Article
-
- You have access
- HTML
- Export citation
Neural effects of a short-term virtual reality self-training program to reduce social anxiety
-
- Journal:
- Psychological Medicine / Volume 52 / Issue 7 / May 2022
- Published online by Cambridge University Press:
- 03 September 2020, pp. 1296-1305
-
- Article
- Export citation
