In this paper, we prove the global exstence of weak solutions for a porous medium dynamics of m species moving between two domains separated by a zero-thickness membrane. On this membrane, Kedem–Katchalsky conditions are considered, and the study is characterized by natural structural conditions applied to the nonlinear reactive terms. The global existence is established under the assumption that these reactive terms are bounded in  $L^1$. This problem has already been analyzed in the linear diffusion case by Ciavolella and Perthame in Ciavolella and Perthame (2021, Journal of Evolution Equations 21, 1513–1540). The present work constitutes an extension for nonlinear diffusion, particularly of the porous medium type, in the form
$L^1$. This problem has already been analyzed in the linear diffusion case by Ciavolella and Perthame in Ciavolella and Perthame (2021, Journal of Evolution Equations 21, 1513–1540). The present work constitutes an extension for nonlinear diffusion, particularly of the porous medium type, in the form  $\partial _t v_i - \Delta v_i^{r_i} = R_i$, for an exponent
$\partial _t v_i - \Delta v_i^{r_i} = R_i$, for an exponent  $r_i < 2$. The case
$r_i < 2$. The case  $r_i \geq 2$ remains an open problem. This paper is an adaptation of the ideas from Ciavolella and Perthame (2021, Journal of Evolution Equations 21, 1513–1540), with new strategies to overcome the appearance of nonlinearity and degeneracy in the diffusion term.
$r_i \geq 2$ remains an open problem. This paper is an adaptation of the ideas from Ciavolella and Perthame (2021, Journal of Evolution Equations 21, 1513–1540), with new strategies to overcome the appearance of nonlinearity and degeneracy in the diffusion term.
We study the Cauchy problem on the real line for the nonlocal Fisher-KPP equation in one spatial dimension,where  \begin{equation*} u_t = D u_{xx} + u(1-\phi *u), \end{equation*}
\begin{equation*} u_t = D u_{xx} + u(1-\phi *u), \end{equation*} $\phi *u$ is a spatial convolution with the top hat kernel,
$\phi *u$ is a spatial convolution with the top hat kernel,  $\phi (y) \equiv H\left (\frac{1}{4}-y^2\right )$. After observing that the problem is globally well-posed, we demonstrate that positive, spatially periodic solutions bifurcate from the spatially uniform steady state solution
$\phi (y) \equiv H\left (\frac{1}{4}-y^2\right )$. After observing that the problem is globally well-posed, we demonstrate that positive, spatially periodic solutions bifurcate from the spatially uniform steady state solution  $u=1$ as the diffusivity,
$u=1$ as the diffusivity,  $D$, decreases through
$D$, decreases through  $\Delta _1 \approx 0.00297$ (the exact value is determined in Section 3). We explicitly construct these spatially periodic solutions as uniformly valid asymptotic approximations for
$\Delta _1 \approx 0.00297$ (the exact value is determined in Section 3). We explicitly construct these spatially periodic solutions as uniformly valid asymptotic approximations for  $D \ll 1$, over one wavelength, via the method of matched asymptotic expansions. These consist, at leading order, of regularly spaced, compactly supported regions with width of
$D \ll 1$, over one wavelength, via the method of matched asymptotic expansions. These consist, at leading order, of regularly spaced, compactly supported regions with width of  $O(1)$ where
$O(1)$ where  $u=O(1)$, separated by regions where
$u=O(1)$, separated by regions where  $u$ is exponentially small at leading order as
$u$ is exponentially small at leading order as  $D \to 0^+$. From numerical solutions, we find that for
$D \to 0^+$. From numerical solutions, we find that for  $D \geq \Delta _1$, permanent form travelling waves, with minimum wavespeed,
$D \geq \Delta _1$, permanent form travelling waves, with minimum wavespeed,  $2 \sqrt{D}$, are generated, whilst for
$2 \sqrt{D}$, are generated, whilst for  $0 \lt D \lt \Delta _1$, the wavefronts generated separate the regions where
$0 \lt D \lt \Delta _1$, the wavefronts generated separate the regions where  $u=0$ from a region where a steady periodic solution is created via a distinct periodic shedding mechanism acting immediately to the rear of the advancing front, with this mechanism becoming more pronounced with decreasing
$u=0$ from a region where a steady periodic solution is created via a distinct periodic shedding mechanism acting immediately to the rear of the advancing front, with this mechanism becoming more pronounced with decreasing  $D$. The structure of these transitional travelling wave forms is examined in some detail.
$D$. The structure of these transitional travelling wave forms is examined in some detail.
In a smoothly bounded domain  $\Omega \subset \mathbb{R}^n$,
$\Omega \subset \mathbb{R}^n$,  $n\ge 1$, this manuscript considers the homogeneous Neumann boundary problem for the chemotaxis systemwith parameter
$n\ge 1$, this manuscript considers the homogeneous Neumann boundary problem for the chemotaxis systemwith parameter  \begin{eqnarray*} \left \{ \begin{array}{l} u_t = \Delta u - \nabla \cdot (u\nabla v), \\[5pt] v_t = \Delta v + u - \alpha uv, \end{array} \right . \end{eqnarray*}
\begin{eqnarray*} \left \{ \begin{array}{l} u_t = \Delta u - \nabla \cdot (u\nabla v), \\[5pt] v_t = \Delta v + u - \alpha uv, \end{array} \right . \end{eqnarray*} $\alpha \gt 0$ and with coincident production and uptake of attractants, as recently emphasized by Dallaston et al. as relevant for the understanding of T-cell dynamics.
$\alpha \gt 0$ and with coincident production and uptake of attractants, as recently emphasized by Dallaston et al. as relevant for the understanding of T-cell dynamics.
It is shown that there exists  $\delta _\star =\delta _\star (n)\gt 0$ such that for any given
$\delta _\star =\delta _\star (n)\gt 0$ such that for any given  $\alpha \ge \frac{1}{\delta _\star }$ and for any suitably regular initial data satisfying
$\alpha \ge \frac{1}{\delta _\star }$ and for any suitably regular initial data satisfying  $v(\cdot, 0)\le \delta _\star$, this problem admits a unique classical solution that stabilizes to the constant equilibrium
$v(\cdot, 0)\le \delta _\star$, this problem admits a unique classical solution that stabilizes to the constant equilibrium  $(\frac{1}{|\Omega |}\int _\Omega u(\cdot, 0), \, \frac{1}{\alpha })$ in the large time limit.
$(\frac{1}{|\Omega |}\int _\Omega u(\cdot, 0), \, \frac{1}{\alpha })$ in the large time limit.
We establish asymptotic formulas for all the eigenvalues of the linearization problem of the Neumann problem for the scalar field equation in a finite interval\[ \begin{cases} \varepsilon^2u_{xx}-u+u^3=0, & 0< x<1,\\ u_x(0)=u_x(1)=0. \end{cases} \]In the previous paper of the third author [T. Wakasa and S. Yotsutani, J. Differ. Equ. 258 (2015), 3960–4006] asymptotic formulas for the Allen–Cahn case $\varepsilon ^2u_{xx}+u-u^3=0$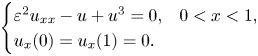
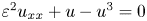 were established. In this paper, we apply the method developed in the previous paper to our case. We show that all the eigenvalues can be classified into three categories, i.e., near $-3$
were established. In this paper, we apply the method developed in the previous paper to our case. We show that all the eigenvalues can be classified into three categories, i.e., near $-3$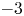 eigenvalues, near $0$
eigenvalues, near $0$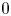 eigenvalues and the other eigenvalues. We see that the number of the near $-3$
eigenvalues and the other eigenvalues. We see that the number of the near $-3$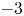 eigenvalues (resp. the near $0$
eigenvalues (resp. the near $0$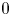 eigenvalues) is equal to the number of the interior and boundary peaks (resp. the interior peaks) of a solution for the nonlinear problem. The main technical tools are various asymptotic formulas for complete elliptic integrals.
eigenvalues) is equal to the number of the interior and boundary peaks (resp. the interior peaks) of a solution for the nonlinear problem. The main technical tools are various asymptotic formulas for complete elliptic integrals.
Exact solutions are constructed for a class of nonlinear hyperbolic reaction-diffusion equations in two-space dimensions. Reduction of variables and subsequent solutions follow from a special nonclassical symmetry that uncovers a conditionally integrable system, equivalent to the linear Helmholtz equation. The hyperbolicity is commonly associated with a speed limit due to a delay, 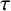 $\tau $, between gradients and fluxes. With lethal boundary conditions on a circular domain wherein a species population exhibits logistic growth of Fisher–KPP type with equal time lag, the critical domain size for avoidance of extinction does not depend on
$\tau $, between gradients and fluxes. With lethal boundary conditions on a circular domain wherein a species population exhibits logistic growth of Fisher–KPP type with equal time lag, the critical domain size for avoidance of extinction does not depend on 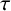 $\tau $. A diminishing exact solution within a circular domain is also constructed, when the reaction represents a weak Allee effect of Huxley type. For a combustion reaction of Arrhenius type, the only known exact solution that is finite but unbounded is extended to allow for a positive
$\tau $. A diminishing exact solution within a circular domain is also constructed, when the reaction represents a weak Allee effect of Huxley type. For a combustion reaction of Arrhenius type, the only known exact solution that is finite but unbounded is extended to allow for a positive 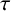 $\tau $.
$\tau $.

