


Non-technical Summary
Ophiuroids (brittle stars) have been major components of marine seafloor communities since the early Paleozoic (about 450 million years ago). Past paleontological studies, especially those dealing with Paleozoic specimens, have based taxonomic descriptions on whole skeletons while mostly overlooking isolated parts, causing a large gap in our knowledge of Paleozoic ophiuroids. Recent studies of Mesozoic and Cenozoic ophiuroids, however, have examined the fine-scale morphological details of ophiuroid arm plates and have documented useful characters for taxonomic assignment. Here, we use similar methods for examining disarticulated ophiuroids to describe a Late Mississippian (Serpukhovian) ophiuroid fauna based solely on dissociated ossicles, preserved as microfossils and sieved from shale samples collected from the Indian Springs Shale Member of the Big Clifty Formation in Sulphur, Indiana, USA. We describe 11 species in total, 10 of which are new to science. The assemblage described in this paper significantly increases the known ophiuroid diversity in the Mississippian, yielding more species than all previous reports on Mississippian ophiuroids combined. Furthermore, our study shows that the evolution of the modern ophiuroid clade began much earlier than expected. Our results imply that the microfossil record of ophiuroids is paramount to unveiling the true paleobiodiversity of this evolutionarily important echinoderm group.
Introduction
Ophiuroids, commonly referred to as brittle stars, are a significant component of modern and ancient seafloor communities. They are extremely widespread and occupy most marine ecosystems from the tropics to the poles and the tidal zones to the deep trenches (Stöhr, Reference Stöhr2012; Stöhr et al., Reference Stöhr, O’Hara and Thuy2012). With approximately 2,100 living species (O’Hara et al., Reference O’Hara, Hugall, Thuy, Stöhr and Martynov2017), they are the most species-rich class of echinoderms and at times are among the most numerous faunal elements within marine ecosystems (Gage and Tyler, Reference Gage and Tyler1991; Aronson et al., Reference Aronson, Thatje, Clarke, Peck, Blake, Wilga and Seibel2007). However, despite their ecological importance, their evolutionary history and diversity patterns remain insufficiently understood (Stöhr Reference Stöhr2012; O’Hara et al., Reference O’Hara, Hugall, Thuy, Stöhr and Martynov2017; Thuy and Stöhr, Reference Thuy and Stöhr2016).
The Late Paleozoic (Mississippian–end Permian) fossil record of ophiuroids is poorly documented globally. Previously published reports of Late Mississippian ophiuroids include approximately nine genera across multiple basins (Harper and Morris, Reference Harper and Morris1978; Jell, Reference Jell1997; Hunter and McNamara, Reference Hunter and McNamara2017; Blake and Nestell, Reference Blake and Nestell2019; Harris et al., Reference Harris, Ettensohn and Carnahan-Jarvis2020). Because these reports are based almost exclusively on relatively rare finds of articulated skeletal material, they under-represent the diversity in the fauna and are biased to the preservation of larger, more robust species over small, fragile ones.
Unlike Paleozoic ophiuroid taxonomy, which relies heavily on the morphology of fully articulated skeletons, recent studies have significantly expanded the known Mesozoic and Cenozoic ophiuroid paleobiodiversity by including taxonomically informative disarticulated skeletal plates (e.g., Jagt, Reference Jagt2000; Thuy, Reference Thuy2013; Thuy and Numberger-Thuy, Reference Thuy and Numberger-Thuy2021). Knowledge on the taxonomic value of ophiuroid ossicles was significantly advanced by the studies of Hess (Reference Hess1962) and Thuy and Stöhr (Reference Thuy and Stöhr2011), who detailed micromorphologies of the ophiuroid lateral arm plates and how they can be used for the assignment of genera and species. However, these studies focused solely on extant clades, ignoring the extinct Paleozoic clades whose ossicle morphologies often differ fundamentally from those of the extant clades (e.g., Hotchkiss and Glass, Reference Hotchkiss and Glass2012). Currently, there is little systematic framework for using ophiuroid lateral arm plates and other ossicles to identify extinct Paleozoic clades (but see Boczarowski, Reference Boczarowski2001; Thuy et al., Reference Thuy, Eriksson, Kutscher, Lindgren, Numberger-Thuy and Wright2022, Reference Thuy, Knox, Numberger-Thuy, Smith and Sumrall2023). As a result, Paleozoic ophiuroid diversity has been poorly documented globally except for a few notable Lagerstätten with exceptional preservation of intact skeletons (Jell, Reference Jell1997; Glass, Reference Glass2006) and few studies using dissociated skeletal plates (Boczarowski, Reference Boczarowski2001; Thuy et al., Reference Thuy, Eriksson, Kutscher, Lindgren, Numberger-Thuy and Wright2022).
Here, we use the ossicle identification methods successfully established for Mesozoic and Cenozoic ophiuroids and extrapolate them to a newly collected assemblage of well-preserved ophiuroid ossicles from the Upper Mississippian (Serpukhovian) Indian Springs Shale Member of the Big Clifty Formation of Crawford Co., Indiana, USA. The material documents a rich and diverse fauna of fossil brittle stars collected as isolated ossicles in washed sediment residues. Analyses of the collected ossicles show surprisingly high taxonomic diversity—higher than the previously known global ophiuroid diversity from the Mississippian (Spencer and Wright, Reference Spencer, Wright and Moore1966; Sprinkle, Reference Sprinkle, Broadhead and Waters1980; Chen and McNamara, Reference Chen and McNamara2005; Hunter and McNamara, Reference Hunter and McNamara2017)—indicating a major sampling bias. Among these isolated ossicles, ophiuroid arm plates, especially lateral arm plates (plates that hold spine articulations and cover the sides of the arm) and, to a lesser extent, the ambulacrals or vertebrae when fused in pairs (ossicles that core the arm) have enough morphological features to determine specific and generic assignments.
The aim of the present paper is to systematically describe the ophiuroid faunal composition of the Indian Springs Shale Member and discuss its implications for late Paleozoic ophiuroid diversity.
Locality and geologic setting
Samples used in this study were collected from a several meters thick, light-gray, fossiliferous shale interbedded with hard limestone layers, referred to as the Indian Springs Shale Member of the Big Clifty Formation (Horowitz et al., Reference Horowitz, Mamet, Neves, Potter and Rexroad1979; Kelly, Reference Kelly1984; Nelson et al., Reference Nelson, Smith, Tregworgy, Keith and Furer2002). The Indian Springs Shale Member specifically includes a lower mudstone interval and an upper shale and limestone interval (Shaver et al., Reference Shaver, Ault, Burger, Carr and Droste1986; Horowitz and Kelly, Reference Horowitz, Kelly and Biggs1987), which contacts the overlying Haney Limestone by a locally minor disconformity (Nelson et al., Reference Nelson, Smith, Tregworgy, Keith and Furer2002). The shale was collected from exposures in southeastern Indiana at the intersection of Interstate 64 and Main Street in Crawford County, Sulphur, Indiana (38°14′33″N, 86°28′09″W; Fig. 1). Approximately 5 kilograms of sediment were collected and processed from the shale layer exposed along the roadside.

Figure 1. Locality map showing the location of the Upper Mississippian (Serpukhovian) Indian Springs Shale Member of the Big Clifty Formation of Crawford Co., Indiana, USA. Friable shale was collected from a cut (marked with a star) on the west entrance ramp of westbound I-64 where it intersects State Route 237.
The locality is Upper Mississippian (Serpukhovian) and represents a tidally influenced, shallow water ramp with clastic sediment influx from deltas to the east-northeast (Klein, Reference Klein1977; Nelson et al., Reference Nelson, Smith, Tregworgy, Keith and Furer2002). The shale is fossil rich and yields brachiopods, echinoderms, bryozoans, and fish teeth (Smith and Sumrall, Reference Smith and Sumrall2019). Echinoderms (mostly identified from disarticulated ossicles) are diverse and abundant, including crinoids (in particular, Pterotocrinus), blastoids (Pentremites), edrioasteroids, echinoids, ophiuroids, and holothuroids (Shaver et al., Reference Shaver, Ault, Burger, Carr and Droste1986; Jell, Reference Jell1997; Nelson et al., Reference Nelson, Smith, Tregworgy, Keith and Furer2002; Smith and Sumrall, Reference Smith and Sumrall2019).
Specimens in this study were collected from traditional lands of the Myaamia, Kaskaskia, Osage, and Shawandasse Tula peoples.
Materials and methods
The ophiuroid material described in this paper was retrieved from washed, sieved residues of bulk sediment samples. Micropaleontological samples were processed by immersing shale residues overnight in a 3-percent hydrogen peroxide and water bath to disaggregate the shale and separate individual skeletal fragments. Sediment was washed to remove the clay fraction, and the process was repeated until the sediment residue was clean. The dried residue was sieved down to 125 μm and hand-picked using a dissection microscope and a wet paintbrush. Selected ossicles were cleaned using a needle under high magnification, as needed, and a sonic water bath. Cleaned ossicles were mounted on aluminum stubs and coated with silver for scanning electron microscopy.
Lateral arm plates were compared to illustrations and descriptions in publications for the purpose of identification. Material here is described following the terminology and procedures of Thuy and Stöhr (Reference Thuy and Stöhr2011) and Gladwell (Reference Gladwell2018). Higher-level classification follows O’Hara et al. (Reference O’Hara, Stöhr, Hugall, Thuy and Martynov2018) for the modern taxa and Gladwell (Reference Gladwell2018) for the Paleozoic ones.
Repository and institutional abbreviation
All ophiuroid material described here is reposited in the collections of the Cincinnati Museum Center with the CMCIP prefix.
Systematic paleontology
Subphylum Asterozoa Zittel, Reference Zittel and von1895
Class Ophiuroidea Gray, Reference Gray1840
Order Stenurida Spencer, Reference Spencer1951
Suborder Parophiurina Jaekel, Reference Jaekel1923
Family Palaeuridae Spencer, Reference Spencer1951
Genus Umerophiura Boczarowski, Reference Boczarowski2001
Type species
Umerophiura opiparia Boczarowski, Reference Boczarowski2001. From the Late Devonian of Poland.
Umerophiura daki new species

Figure 2. SEM photographs of lateral arm plates; arrows on external views indicate distal direction. (1–10) Umerophiura daki n. sp., scale bar = 500 μm. (1) Paratype CMCIP-94540, proximal/median LAP, exterior view; (2, 3, 5) paratype CMCIP-94543, median/distal LAP, interior, exterior and ventral views, note the smooth main plate body, reticulated stereom on the articulation peg, ventral fossa on the exterior surface and the coarse stereom and well-defined podial basin on the interior surface; small spine articulations ring the plate body on the interior surface; (4, 8, 9) holotype CMCIP-94541, proximal LAP, dorsal, exterior and interior views; (6, 7, 10) paratype CMCIP-94542, unusual proximal LAP, interior, dorsal and exterior views. (11–27) Strataster lisae n. sp., scale bar = 500 μm except (26) = 250 μm. (11, 12) Paratype CMCIP-94594, median LAP, exterior and interior views; (13, 14) paratype CMCIP-94596, distal LAP, interior and exterior views; (15, 16) paratype CMCIP-94591, median LAP, interior and exterior views; (17, 22–24) paratype CMCIP-94593, proximal LAP, interior, exterior, dorsal and ventral views; (20, 21, 25–27) holotype CMCIP-94592, proximal left LAP, dorsal, exterior, ventral, detail of spine base and interior views. Note the pit-like spine bases along the distal margin.
Holotype
Holotype specimen CMCIP-94541 (Fig. 2.4, 2.8, 2.9) is a proximal left lateral arm plate; Crawford County, Indiana, USA; Indian Springs Shale Member, Big Clifty Formation (Serpukhovian).
Paratypes
Paratypes include CMCIP-94540, 94542, 94543; Crawford County, Indiana, USA; Indian Springs Shale Member, Big Clifty Formation (Serpukhovian).
Diagnosis
Species of Umerophiura with racket-shaped lateral arm plates bearing short digitate processes on the distal margin and short, rounded, peg-like vertebral articulation. Outer surface smooth, devoid of ornamentation. Inner surface mostly smooth with slightly coarse tuberculation along distal margin. Distal edge with spiny, flattened triangle-shaped protrusions and lined underneath by medium-sized spine articulations composed of shallow depressions with central peg. Inner surface with large, deep podial basin.
Occurrence
Umerophiura daki n. sp. is known from the Upper Mississippian (Serpukhovian) Indian Springs Shale Member of the Big Clifty Formation, Crawford County, Indiana, USA.
Description of holotype
CMCIP-94541 (Fig. 2.4, 2.8, 2.9), dissociated proximal lateral arm plate, slightly longer than high, with racket-like outline; main body of plate shaped like glove, elliptical with long axis extending from dorso-proximal to central-ventral, with strongly convex distal edge; peg-like proximal portion extending from ventro-proximal margin of plate body; dorsal edge of peg convex, ventral edge concave; proximal part of peg wider than distal part; proximal tip of peg with central depression dorsally and ventrally bordered by horizontal ridges; plate body convex when viewed axially from dorsal, ventral, and distal sides (Fig. 2.4); articulation peg cylindrical in shape when viewed axially, with a slight constriction upon entering the main plate body; stereom on outer surface of main plate body finely meshed and devoid of tuberculation (Fig. 2.8); stereom of proximal peg more coarsely meshed, with large, irregular, reticulum-like depressions upon entering main plate body; distal edge of main plate body with deep furrow; up to nine rounded, flattened, triangular protrusions emerging from within furrow, largest in middle and decreasing in size ventrally and dorsally; spine articulations just below triangular protrusions, composed of medium-sized depressions with a tiny central knob.
Inner side of lateral arm plate (Fig. 2.9) broadly concave with coarsely meshed, reticulate stereom and with large, central depression ventral with large, widely concave but shallow tentacle notch distally bordered by a thickened, knob-like ventro-distal plate edge.
Description and variation of paratypes
Paratype CMCIP-94540 (Fig. 2.1), proximal/median lateral arm plate, plate slightly longer than high, similar to holotype but with kink in proximal peg; stereom on outer surface of peg more coarsely meshed than on outer surface of main plate body; distal edge bordering furrow not as sharp and regular as in holotype; spine articulations and inner side as in holotype.
Paratype CMCIP-94542 (Fig. 2.6, 2.7, 2.10) dissociated proximal lateral arm plate, nearly as high as long, generally similar to holotype but with more strongly ellipsoid main plate body; proximal peg with kink as in paratype CMCIP-94540; outer surface with more coarsely meshed stereom on peg than on main plate body; spine articulations and inner side as in holotype.
Paratype CMCIP-94543 (Fig. 2.2, 2.3, 2.5) dissociated median to distal lateral arm plate, at least 1.5 times longer than high; up to seven triangular protrusions with spine articulations similar to those of holotype in furrow along distal plate edge; inner side as in holotype.
Etymology
This species is named in honor of Dak, the pet doxie of NSS, which provided comfort during the writing of this manuscript.
Remarks
The lateral arm plates of Umerophiura daki n. sp. are characterized by their broad, glove-shaped main plate body, proximal rounded peg-like vertebral articulation, distal edge lined with flattened triangular fixed spinelets, and a depression on the inner surface leading to a prominent incision for the podial basin and opening. This species most closely resembles isolated lateral arm plates (LAPs) from the Devonian of Poland described by Boczarowski (Reference Boczarowski2001) as Umerophiura sp., especially the following characters: “raquet-shaped” main body plate with prominent club like articulation peg protruding from the proximal margin, wedge-shaped fixed spinelets aligning the distal margin, finely meshed surface stereom, and a large tentacle notch on the ventral edge of the plate. Umerophiura daki n. sp. differs from U. opiparia by having a glove-like main plate and rounded peg-like proximal vertebral articulation. Umerophiura umera Boczarowski, Reference Boczarowski2001, shows the strongest similarity but differs with respect to the concavity of the main plate body, the angle between the peg and the main plate body, and the more prominent and rounded articulation peg.
Boczarowski (Reference Boczarowski2001) assigned Umerophiura to the parophiurin family Palaeuridae without providing arguments in support of this classification. Unfortunately, virtually no attention has been paid to the morphology of lateral arm plates in stenuroid taxa, except perhaps for the general outline of the plates in ventral and/or dorsal views. Furthermore, published illustrations generally fail to show details of the lateral arm plates. As a result, comparison between the previously published articulated skeletons and the dissociated lateral arm plates of Boczarowski (Reference Boczarowski2001) and of the present material is difficult. Pending a more thorough assessment of the lateral arm plate morphologies of the currently known stenuroid taxa, we conform with Boczarowski (Reference Boczarowski2001) and treat the material described herein as an early Carboniferous record of the palaeurid genus Umerophiura.
Order Oegophiurida Matsumoto, Reference Matsumoto1915
Suborder Lysophiurina Gregory, Reference Gregory1896
Family Protasteridae Miller, Reference Miller1889
Genus Strataster Kesling and Le Vasseur, Reference Kesling and Le Vasseur1971
Type species
Strataster ohioensis Kesling and Vasseur, Reference Kesling and Le Vasseur1971. From the Early Mississippian of Ohio.
Strataster lisae new species
Holotype
Specimen CMCIP-94592 is a proximal left arm plate; Crawford County, Indiana, USA; Indian Springs Shale Member, Big Clifty Formation (Serpukhovian).
Paratypes
Paratypes include CMCIP-94591 and 94593–94596; Crawford County, Indiana, USA; Indian Springs Shale Member, Big Clifty Formation (Serpukhovian).
Diagnosis
Species of Strataster with trapezoidal-shaped lateral arm plates with up to three relatively small lateral arm spine articulations close to distal edge, and row of small, inconspicuous ambulacral groove spine articulations along ventral plate edge.
Occurrence
Strataster lisae n. sp. is known from the Upper Mississippian (Serpukhovian) Indian Springs Shale Member of the Big Clifty Formation, Crawford County, Indiana, USA.
Description of holotype
Proximal left lateral arm plate CMCIP-94592 (Fig. 2.20, 2.21, 2.25–2.27), slightly longer than high with rounded trapezoidal outline, highest distally, tapering slightly towards proximal edge; dorsal edge straight to slightly concave; ventral edge oblique and convex, outline devoid of incisions or protuberances; distal edge convex; proximal edge slightly convex; outer surface with ornamentation consisting of fine tuberculation covering entire surface; proximal edge of lateral arm plate with large, poorly defined depression bordered dorsally and ventrally by a small, poorly defined, inconspicuous but prominent knobs; distal plate margin with three large spine articulations at same level of outer surface in vertical, slightly oblique row approximately 50 microns from distal edge of lateral arm plate; size of spine articulations and distance between spine articulations increasing towards dorsal edge (i.e., dorsalmost spine articulation largest); each spine articulation composed of single, large opening encompassed by two comma-shaped, opposed, vertical ridges. Internal side of lateral arm plate with a large, highly elevated and well-defined vertebral articular knob with raised margins shaped like a belt buckle, and weakly depressed center; knob close to proximal ventral margin and distally bordered by a deep vertical depression (podial basin) dorsally enclosed by dorsal edge of the lateral arm plate and ventrally open; ventral edge of lateral arm plate around opening of central depression lined by small, inconspicuous groove spine articulations consisting of weakly raised cones with a tiny central opening; distal edge of lateral arm plate smooth and slightly pinnacled, devoid of spurs.
Description and variation of paratypes
Median lateral arm plate CMCIP-94591 (Fig. 2.15, 2.16), more elongate, longer than high, with rounded rectangular outline; ventral edge less strongly oblique than in holotype (Fig. 2.27); outer surface ornamentation as in holotype; three spurs on proximal edge but central one closer to the dorsal one; three spine articulations but without size gradient; interior side and axial views same as in holotype.
Distal lateral arm plate CMCIP-94596 (Fig. 2.13, 2.14), more rounded overall with concave proximal margin; with only central and ventral spurs visible; outer surface ornamentation as in holotype but with groove spine articulations poorly visible, axial views same as in holotype.
Etymology
This species is named in honor of Lis Thuy.
Remarks
The lateral arm plates described above are characterized by their flat shape and trapezoidal plate outline, the relatively large lateral spine articulations composed of a single opening encompassed by ridges and just inset from the distal margin, a row of ambulacral groove spine articulations along the ventral edge, and a vertebral articular knob on the proximal ventral inner surface that resembles a belt buckle. These are characters typically seen in protasterid ophiuroids, which is why we assign the present material to the family Protasteridae. While currently known protasterid lateral arm plates tend to share a common general morphology, it is noteworthy that some genera seem to have different morphology to lateral spine articulations. Strataster ohioensis Kessling and Le Vasseur, Reference Kesling and Le Vasseur1971, Strataster stuckenbergi (Rilett, Reference Rilett1971), and Strataster maciverorum Hotchkiss, Reference Hotchkiss1993, all have the single opening of their lateral arm spine articulation encompassed by two vertical ridges. In contrast, Eugasterella logani (Hall and Whitney, Reference Hall and Whitney1858), Eugasterella thorni Kesling, Reference Kesling1969, and Eugasterella africana Jell and Theron, Reference Jell and Theron1999, all have simple openings encompassed by horizontal ridges. Remarkably, the specimens originally described as Strataster devonicus Kesling, Reference Kesling1972, have lateral arm spine articulations similar to those of Eugasterella (i.e. simple openings encompassed by horizontal ridges), and were subsequently transferred to the latter genus by Hotchkiss (Reference Hotchkiss1993), albeit because of other characters. This corroborates that genus-level taxa tend to share similar lateral arm plate morphologies, implying that dissociated lateral arm plates can indeed be used for taxonomic identifications even in previously neglected Paleozoic assemblages.
Because of the lateral arm spine articulation shape, we assign the material described above to the genus Strataster. They differ from the previously known species of that genus in having smaller lateral arm spine articulations and inconspicuous ambulacral groove spine articulations.
Genus Schoenaster Meek and Worthen, Reference Meek and Worthen1860
Type species
Schoenaster fimbriatus Meek and Worthen, Reference Meek and Worthen1860. From the Carboniferous of Illinois.
Schoenaster limbeckae new species

Figure 3. Schoenaster limbeckae n. sp.; SEM photographs of lateral arm plates; scale bar = 500 μm except (27, 35) = 250 μm; arrows on external views indicate distal direction. (1–4) Paratype CMCIP-94553, median right LAP in ventral, exterior, dorsal and interior views; (5, 6, 9, 13) paratype CMCIP-94555, median right LAP in ventral, dorsal, exterior and interior views; (7, 8, 12, 16) paratype CMCIP-94558, median right LAP in ventral, exterior, interior and dorsal views; (10, 11, 14, 15) paratype CMCIP-94561, median right LAP in dorsal, interior, exterior and ventral views; (17–20) paratype CMCIP-94557, distal right LAP in interior, exterior, dorsal and ventral views; (21–25, 35) paratype CMCIP-94554, very proximal right LAP, possibly in disk in oblique distal, exterior, dorsal, interior, ventral and detail of lateral spine articulation views; (26–30) holotype CMCIP-94552, proximal right LAP in interior, detail of groove spine articulations, exterior, ventral and dorsal views; (31–34) paratype CMCIP-94559, distal right LAP in exterior, interior, dorsal and ventral views.
Holotype
Specimen CMCIP-94552 (Fig. 3.26–3.30) is a proximal right lateral arm plate; Crawford County, Indiana, USA; Indian Springs Shale Member, Big Clifty Formation (Serpukhovian).
Paratypes
Paratypes include CMCIP-94553–94559; Crawford County, Indiana, USA; Indian Springs Shale Member, Big Clifty Formation (Serpukhovian).
Diagnosis
Species of Schoenaster with lateral arm plates of oval outline; strongly convex ventral edge with a row of conspicuous ambulacral groove spine articulations; two to three large lateral arm spine articulations in a strongly oblique row distally bordered by a narrow, pinnacled plate edge, each spine articulation composed of a single opening encompassed by two shifted (slightly offset from the other), comma-shaped ridges; outer proximal edge of the lateral arm plate with a central depression bordered by central and dorsal spur.
Occurrence
Schoenaster limbeckae n. sp. is known from the Upper Mississippian (Serpukhovian) Indian Springs Shale Member of the Big Clifty Formation, Crawford County, Indiana, USA.
Description of holotype
Proximal right lateral arm plate CMCIP-94552 (Fig. 3.26–3.30), slightly longer than high, of oval outline, with rounded dorsal, distal and ventral edges, wavy proximal edge; plate bulky and thick but not arched; outer surface bulging, covered by moderately coarsely meshed stereom devoid of tubercles; outer proximal edge pinnacled, with poorly defined, semi-circular central depression and two large, round, moderately well-defined, prominent but not protruding central spurs; two large lateral spine articulations in strongly oblique row along the ventro-distal edge, sunken in shallow, distalwards oriented depressions, separated from plate edge by thinned ledge; lateral spine articulations composed of single large opening encompassed by two comma-shaped, shifted, vertical ridges; ventral edge of plate strongly convex, lined by row of six protruding ambulacral groove spine articulations that lie adradial to thin narrow and shallow groove along ventral margin. Internal side of lateral arm plate with large, poorly defined, weakly prominent, round vertebral articular ridge close to dorso-proximal corner of plate, showing raised margins and slightly sunken center, resulting in general belt-buckle shape; inner distal edge of plate corresponding to ledge bordering lateral spine articulations, offset from remaining plate and with rounded, slightly thickened dorso- and ventro-distal edges; no spur discernible on inner distal edge; central depression large but shallow, covered by more coarsely meshed stereom, ventrally bordered by row of cone-shaped ambulacral groove spine articulations (Fig. 3.27).
Description and variation of paratypes
Proximal lateral arm plate CMCIP-94554 (Fig. 3.21–3.25, 3.35) probably from within disc, slightly longer than holotype and with a more angular outline; outer surface stereom and spur on outer proximal edge as in holotype; two large lateral spine articulations similar to those of holotype (Fig. 3.28); ledge distally bordering lateral spine articulations larger, with a conspicuous, pointed ventral tip; ventral edge of lateral arm plate weakly convex, with smaller ambulacral groove spine articulations; internal side as in holotype but with vertebral articular knob in a more central position.
Median lateral arm plates CMCIP-94553 (Fig. 3.1–3.4), CMCIP-94555 (Fig. 3.5, 3.6, 3.9, 3.13), CMCIP-94561 (Fig. 3.10, 3.11, 3.14, 3.15), and CMCIP-94558 (Fig. 3.7, 3.8, 3.12, 3.16) slightly more elongate than holotype, of similarly oval outline and bulky, non-arched shape; outer surface and outer proximal edge with spur as in holotype; three lateral spine articulations similar to those of holotype, in a strongly oblique row along the ventro-distal plate edge; ventral edge of plate strongly convex, with up to five ambulacral groove spine articulations similar to those of holotype; internal side as in holotype.
Distal lateral arm plates CMCIP-94557 (Fig. 3.17–3.20), CMCIP-94559 (Fig. 3.31–3.34) clearly longer than high, of rounded rectangular to rounded trapezoidal outline; generally similar to holotype, with two lateral spine articulations; convex ventral edge with four to five ambulacral groove spine articulations; internal side as in holotype.
Etymology
This species is named in honor of Maggie Limbeck who helped with the preparation of this manuscript.
Remarks
The lateral arm plates described above show striking similarities with the lateral arm plates of the Schoenaster fimbriatus (Meek and Worthen, Reference Meek and Worthen1860) specimens figured and described by Jell (Reference Jell1997). In particular the rounded, bulging shape, the size, position and number of lateral arm spine articulations, the presence of several relatively large ambulacral groove spine articulations, and the adradially pointing proximal tip of the lateral arm plates support the assignment to Schoenaster. The genus was redescribed in detail by Jell (Reference Jell1997) who assigned it to the family Encrinasteridae based on the wide lateral arm plates. The presence of well-developed ambulacral groove spine articulations, however, precludes encrinasterid affinities (Harper and Morris, Reference Harper and Morris1978; Reddy et al., Reference Reddy, Thuy, Reid and Gess2023). We therefore suggest transferring Schoenaster to the family Protasteridae whose members are characterized by their large and well-developed lateral and ambulacral groove spine articulations. The shape of the ventral surface of the ambulacrals and the dorsal plating of the disc and the arms also align with the reassignment to the Protasteridae.
Schoenaster seems to be common and widespread in the Mississippian of the United States (Harris et al., Reference Harris, Ettensohn and Carnahan-Jarvis2020). Four species are currently assigned to the genus Schoenaster (Harris et al., Reference Harris, Ettensohn and Carnahan-Jarvis2020), ranging from the Kinderhookian (Tournaisian) to the Chesterian (Serpukhovian). Apart from the type species S. fimbriatus, however, no currently known species are described in a way that would allow for a detailed comparison with the material presented herein. At least the type specimens of Schoenaster legrandensis Miller and Gurley, Reference Miller and Gurley1888, S. montanus Raymond, Reference Raymond1912, and S. carterensis Harris, Ettensohn, and Carnahan-Jarvis, Reference Harris, Ettensohn and Carnahan-Jarvis2020, seem too poorly preserved to allow for a meaningful assessment of the potentially diagnostic characters. The differences between species highlighted by Harris et al. (Reference Harris, Ettensohn and Carnahan-Jarvis2020) likely result from preservation, precluding an unambiguous identification and thus challenging the validity of the respective species. Compared to the lateral arm plates of Schoenaster fimbriatus illustrated by Jell (Reference Jell1997), the specimens described herein are thinner and have larger lateral spine articulations and a finer outer surface stereom. We therefore assign them to the new species Schoenaster limbeckae.
Family Cheiropterasteridae Spencer, Reference Spencer1934
Genus Vandelooaster Jell, Reference Jell1997
Type species
Vandelooaster plicatilis Jell, Reference Jell1997. From the Early Carboniferous of Indiana.
Vandelooaster douglasi new species

Figure 4. SEM photographs of lateral arm plates; scale bars = 500 μm; arrows on external views indicate distal direction. (1–5) Cholaster sp. indet. (1–3) proximal right LAP CMCIP-94550, exterior, distal, and interior views; note the plate is tall and narrow, and the interior appears to lack a vertebral articulation process but is damaged centrally; (4, 5) CMCIP-94551, median to distal LAP, interior and exterior views. (6–26) Vandelooaster douglasi n. sp. (6, 11–13) paratype CMCIP-94546, median LAP, interior, exterior, dorsal, and ventral views; (7–10) paratype CMCIP-94548, distal LAP, interior, dorsal, ventral, and exterior views; (14, 15, 18, 19) holotype CMCIP-94545, proximal right LAP, exterior, interior, dorsal, and ventral views; note the podial basin with tentacle notch and the ventral shelf; (16, 17) paratype CMCIP-94549, proximal LAP, interior and exterior views; (20, 23, 24) paratype CMCIP-94547, somewhat elongate proximal LAP, interior, exterior, and distal views; (21, 22, 25, 26) paratype CMCIP-94544, proximal LAP, exterior, interior, ventral, and dorsal views.
Holotype
Specimen CMCIP-94545 (Fig. 4.14, 4.15, 4.18, 4.19) is a proximal right lateral arm plate; Crawford County, Indiana, USA; Indian Springs Shale Member, Big Clifty Formation (Serpukhovian).
Paratypes
Paratypes include CMCIP-94544, CMCIP-94546–94549; Crawford County, Indiana, USA; Indian Springs Shale Member, Big Clifty Formation (Serpukhovian).
Diagnosis
Species of Vandelooaster with a relatively short, blunt, peg-like proximal plate portion.
Occurrence
Vandelooaster douglasi n. sp. is known from the Upper Mississippian (Serpukhovian) Indian Springs Shale Member of the Big Clifty Formation, Crawford County, Indiana, USA.
Description of holotype
Proximal right lateral plate CMCIP-94545 (Fig. 4.14, 4.15, 4.18, 4.19), wider than high, general spatula-like outline, highest distally, tapering slightly proximally for two-thirds of the plate length; proximal one-third of plate significantly longer than high and with a general peg-like shape; distal edge undulating with slight convexity overall; dorsal edge strongly concave proximally, raising to convex edge distally; acute proximal angle converging proximalwards to form peg-like articulation structure with vertebrae; articulation peg in slightly off-center, more ventral position with respect to the main plate body; main plate body superficially rounded and thick, in dorso-ventral axial views with rounded edge, slightly increasing in height toward plate center, rounded ridge encircling center of the main plate body; distal axial view similar to dorso-ventral view but with rounded ridge along ventro-distal margin; plate rounded in proximal axial view with conical appearance expanding toward plate midline, diminishing toward the plate midline upon meeting the main plate body; outer surface (Fig. 4.14) ornamentation consisting of a very fine tuberculation covering the entire surface, devoid of patterns, with slightly more coarse tuberculation along dorsal edge of flattened, paddle-like main body of plate; slightly raised, vertical ridge in middle of main plate body; distal edge of main plate body lined by a shallow groove, four small, inconspicuous spine articulations directly along the distal edge of plate, slightly protruding as part of distal edge undulation; slight ventralward increase in size of spine articulations and in distance between them; every spine articulation composed of a small, round opening (Fig. 4.18). Interior side of lateral arm plate (Fig. 3.15) with a thick ridge along the dorsal edge of the distal half of the plate; proximal peg bordered by deep depression in center of main body of plate, closed on ventral, distal, and dorsal edges by slight raise in surface of plate, proximal edge of depression opening to flat even surface with rest of plate; small spur consisting of two vertical, ellipsoid, dome-like ridges of equal height and width along the ventral edge of the peg close to main body of plate.
Description and variation of paratypes
Proximal lateral arm plate CMCIP-94547 (Fig. 4.20, 4.23, 4.24) similar to holotype but more elongate and streamlined in outline. Median lateral arm plate CMCIP-94546 (Fig. 4.6, 4.11–4.13) similar to holotype in general outline, but not as wide; four lateral spine articulations as in holotype. Distal lateral arm plate CMCIP-94548 (Fig. 4.7–4.10) similar to holotype but significantly narrower and with much shorter vertebral articular peg; three lateral spine articulations along distal edge.
Etymology
This species is named in honor of Douglas Sumrall.
Remarks
Lateral arm plates of Vandelooaster douglasi n. sp. are characterized by their paddle-like main plate body, spine articulations on a ridge-like extension of the main plate body along the ventro-distal margin, spine articulations composed of simple round openings, and a conical vertebral articulation arising from proximal edge of the main plate body. They most closely resemble the lateral arm plates of the Carboniferous cheiropterasterid Vandelooaster plicatilis Jell, Reference Jell1997. The lateral arm plates (LAPs) described here and those of V. plicatilis both show a flat paddle-like main plate body, spine articulations along a ridge like extension from the ventro-distal margin, and an elongate conical vertebral articulation. Specimens of V. plicatilis viewed both at the National Museum of Natural History (USNM 467196) and those figured by Jell (Reference Jell1997) show that the peg-like proximal portion of the Vandelooaster lateral arm plates described above is shorter and less pointed than in V. plicatilis. We therefore describe the material at hand as a new species, adding a second Mississippian record to the genus Vandelooaster.
Suborder Zeugophiurina Matsumoto, Reference Matsumoto1929
Family Furcasteridae Stürtz, Reference Stürtz1900
Genus Furcaster Stürtz, Reference Stürtz1900
Type species
Furcaster palaeozoicus Stürtz, Reference Stürtz1900. From the Devonian of Germany.
Remarks
The material described in the present paper includes lateral arm plates (LAPs) that show the highly characteristic shape, outer surface ornamentation, vertebral articular knob, and spine articulation morphology of the genus Furcaster (e.g., Haude and Thomas, Reference Haude and Thomas1983; Boczarowski, Reference Boczarowski2001; Gladwell, Reference Gladwell2018). Since this combination of characters is unknown from any other ophiuroid genus, we assign the lateral arm plates in question to Furcaster. The genus is diverse and long-lived, ranging from the Late Ordovician to the Late Devonian (Gladwell, Reference Gladwell2018). From the Carboniferous, only undetermined specimens (Haude, Reference Haude1982) or dissociated vertebrae (Weller, Reference Weller1930) have been described, leaving the post-Devonian record of the genus virtually unknown. The material described herein therefore significantly adds to the fossil record of the genus, contributing the first nominal species from the Carboniferous.
The Furcaster material from Sulphur can be subdivided into three different morphological types. Since the differences among these three types can be followed consistently from proximal to distal LAPs, and since they cannot be explained by ontogenetic variation, we consider the three types as different species.
Furcaster wardi new species

Figure 5. SEM photographs of lateral arm plates; scale bars = 500 μm except (28) = 250 μm; arrows on external views indicate distal direction. (1–31) Furcaster wardi n. sp. (1, 2, 4, 5) paratype CMCIP-94598, distal right LAP, exterior, interior, distal, and ventral views; (6–9) paratype CMCIP-94599, distal left LAP, exterior, ventral, distal, and interior views; (3, 10, 12) paratype CMCIP-94606, median left LAP, distal, interior, and exterior views; (11, 13–15) paratype CMCIP-94604, median right LAP, ventral, distal, interior, and exterior views; (16–19) paratype CMCIP-94603, median left LAP, interior, distal, exterior, and ventral views; (20–22) paratype CMCIP-94605, median left LAP, distal, interior, and exterior views; (23, 29–31) paratype CMCIP-94602, proximal right LAP, distal, ventral, interior, and exterior views; (24–28) holotype CMCIP-94601, proximal right LAP, interior, distal, exterior, ventral, and detail of ventralmost lateral spine articulations views.
Holotype
The holotype specimen of Furcaster wardi n. sp. CMCIP-94601 (Fig. 5.24–5.28) is a proximal right lateral arm plate; Crawford County, Indiana, USA; Indian Springs Shale Member, Big Clifty Formation (Serpukhovian).
Paratypes
Paratypes include CMCIP-94597–94600, CMCIP-94602–94606; Crawford County, Indiana, USA; Indian Springs Shale Member, Big Clifty Formation (Serpukhovian).
Diagnosis
Species of Furcaster with large, relatively robust, very high, strongly arched lateral arm plates; outer surface ornamentation composed of coarsely meshed stereom, with densely spaced pores at a slight angle to the outer surface and arranged in a weak vertical striation; up to eight large lateral spine articulations sunken into notches along distal edge, distally bordered by narrow ledge; up to four ambulacral groove spine articulations sunken in notches along ventral edge.
Occurrence
Furcaster wardi n. sp. is known from the Upper Mississippian (Serpukhovian) Indian Springs Shale Member of the Big Clifty Formation, Crawford County, Indiana, USA.
Description of holotype
Proximal right lateral arm plate CMCIP-94601 (Fig. 5.24–5.28) relatively thick, significantly higher than long; scythe-shaped outline, concave proximally, longest at base and decreasing in length dorsally; ventral portion of plate protruding proximalwards; distal margin evenly convex and rounded; proximal edge concave and lined by band of more finely meshed stereom, well defined and slightly raised ventrally, fading towards dorsal edge of plate, devoid of spurs; outer surface ornamentation composed of coarsely meshed stereom without tubercles but with pores opening at slight distalwards pointing angle with respect to outer surface, densely spaced and arranged in very weak vertical striation; eight large lateral spine articulations sunken in deep notches along distal plate edge, arranged in continuous row distally bordered by a very narrow ledge, with a weak dorsalward increase in size of spine articulations and of the gaps separating them; notches of spine articulations separated by conspicuous, dorso-distalwards pointing protrusions of the outer surface; spine articulations consisting of strongly prominent, lip-shaped vertical ridge in middle of a large pit, proximally merged with a much smaller, lower and thinner, semicircular ridge enclosing a small perforation (Fig. 5.28); ridges of dorsally positioned spine bases parallel to plate surface, ventralmost three becoming increasingly horizontal; ventral edge indented by four ambulacral groove spine articulations similar to lateral spine articulations but smaller, all equal-sized except for a smaller distalmost one; inner side of lateral arm plate with large, thick, strongly prominent, tooth-shaped vertebral articular knob on the ventral portion of the plate, pointing dorsalwards; ventral margin along ambulacral groove spine bases with smooth imperforate stereom (Fig. 5.24).
Description and variation of paratypes
Median lateral arm plate CMCIP-94606 (Fig. 5.3, 5.10, 5.12) nearly as high as long, rounded triangular in outline, less strongly arched, otherwise similar to holotype; six lateral and three ambulacral groove spine articulations, all similar to those of holotype; inner side of lateral arm plate with vertebral articular knob similar to that of holotype but less strongly prominent.
Distal lateral arm plate CMCIP-94598 (Fig. 5.1, 5.2, 5.4, 5.5) slightly longer than high, of rounded trapezoidal outline, very weakly arched; outer surface ornamentation as in holotype; three lateral and two ambulacral groove spine articulations similar to those of holotype; inner side with a large, weakly prominent, ridge-like vertebral articular knob.
Etymology
This species is named in honor of Gabe Ward who provided valuable specimens that helped with the description of this material.
Remarks
The lateral arm plates of Furcaster wardi n. sp. stand out as being considerably larger than those of the other two Furcaster species in the Sulphur assemblage. Specifically, they differ from Furcaster mccantae n. sp. bearing a much higher and strongly arched general shape, the presence of up to eight lateral spine articulations sunken in deep depressions along the distal edge, and the much smaller pores that are at a slight angle to the outer surface and arranged in a weak vertical striation. Compared to the lateral arm plates of Furcaster coulombeae n. sp., those of Furcaster wardi n. sp. are more robust, have the pores of the outer surface stereom more closely spaced and at a higher angle with respect to the outer surface, and have larger lateral spine articulations with a larger perforation and a larger semicircular ridge encompassing the perforation.
Furcaster mccantae new species

Figure 6. SEM photographs of lateral arm plates, scale bars = 250 μm except (16) = 125 μm; arrows on external views indicate distal direction. (1–16) Furcaster mccantae n. sp. (1–3) paratype CMCIP-94610, distal right LAP, exterior, interior, and ventral views; (4–7) paratype CMCIP-94609, median left LAP, exterior, ventral, interior, and distal views; (8, 9, 11, 15, 16) holotype CMCIP-94611, proximal right LAP, distal, exterior, interior, ventral, and detail of lateral spine articulations views; (10, 12–14) paratype CMCIP-94607, proximal right LAP, exterior, interior, distal, and ventral views. (17–25) Furcaster coulombeae n. sp. (17–19, 21, 22) holotype CMCIP-94612, proximal right LAP, dorsal, interior, ventral, distal, and exterior views; (20, 23–25) paratype CMCIP-94613, median to distal right LAP, ventral, interior, dorsal, and exterior views.
Holotype
Holotype specimen CMCIP-94611 (Fig. 6.8, 6.9, 6.11, 6.15, 6.16) is a proximal right lateral arm plate; Crawford County, Indiana, USA; Indian Springs Shale Member, Big Clifty Formation (Serpukhovian).
Paratypes
Paratypes include CMCIP-94607–94610; Crawford County, Indiana, USA; Indian Springs Shale Member, Big Clifty Formation (Serpukhovian).
Diagnosis
Species of Furcaster with moderately large, robust, rounded triangular, low and weakly arched lateral arm plates; outer surface with very coarsely meshed stereom bearing large pores almost perpendicular to outer surface; up to four large lateral spine articulations with very large central perforation, protruding from small, shallow and poorly defined notches along distal plate edge; up to four relatively small ambulacral groove spine articulations along ventral plate edge.
Occurrence
Furcaster mccantae n. sp. is known from the Upper Mississippian (Serpukhovian) Indian Springs Shale Member of the Big Clifty Formation, Crawford County, Indiana, USA.
Description of holotype
Proximal right lateral arm plate CMCIP-94611 (Fig. 6.8, 6.9, 6.11, 6.15, 6.16) as high as long, robust, rounded triangular in outline, weakly arched, with evenly convex distal edge and nearly straight ventral edge; slightly concave proximal edge lined by narrow, poorly defined band of more finely meshed stereom, devoid of spurs or other articular structures; outer surface composed of very coarsely meshed stereom, with large, closely spaced pores nearly perpendicular to outer surface (Fig. 6.9); four large lateral spine articulations in small, shallow, poorly defined notches of distal plate edge, with dorsalward increase in size of notches; spine articulations with very large central opening encompassed by almost circular ridge, surrounded by slightly more finely meshed stereom and separated from neighboring spine articulations by projections of more coarsely meshed outer surface stereom (Fig. 6.16); inner side of lateral arm plate with large, well-defined, weakly prominent, oval vertebral articular knob close to ventro-proximal corner of plate; ventral margin along ambulacral groove spine bases with smooth imperforate stereom; ventral edge with shallow notches of four relatively small, elongate ambulacral groove spine articulations, two median ones larger than distal and proximal ones.
Description and variation of paratypes
Median left lateral arm plate CMCIP-94609 (Fig. 6.4–6.7) longer than high, with rounded trapezoidal outline, with oblique proximal edge lined by relatively wide, poorly defined band of more finely meshed stereom; outer surface as in holotype; four lateral and four ambulacral groove spine articulations as in holotype; inner side with vertebral articular knob similar to that of holotype but less sharply defined.
Distal right lateral arm plate CMCIP-94610 (Fig. 6.1–6.3) almost 1.5 times longer than high, similar to paratype but with three lateral and three ambulacral groove spine articulations; inner side as in paratype.
Etymology
This species is named in honor of Molly McCanta who provided SEM access, greatly assisting this research.
Remarks
Furcaster mccantae n. sp. differs from its Sulphur congeners in having much lower, weakly arched lateral arm plates with very coarse outer surface stereom bearing pores almost perpendicular to the outer plate surface, and only four, rather than eight, large lateral spine articulations with a very large central perforation, protruding from small, shallow and poorly defined notches of the distal plate edge.
Furcaster coulombeae new species
Holotype
Holotype specimen CMCIP-94612 (Fig. 6.17–6.19, 6.21, 6.22) is a proximal right lateral arm plate; Crawford County, Indiana, USA; Indian Springs Shale Member, Big Clifty Formation (Serpukhovian).
Paratype
Paratype CMCIP-94613 (Fig. 6.20, 6.23–6.25); Crawford County, Indiana, USA; Indian Springs Shale Member, Big Clifty Formation (Serpukhovian).
Diagnosis
Species of Furcaster with small, fragile, high and strongly arched, triangular lateral arm plates; outer surface with coarsely meshed stereom with large, widely spaced pores, distalwards pointing at a low angle to outer surface; up to eight small lateral spine articulations sunken in deep, distalwards pointing, equidistant and equally sized notches of the distal plate edge, spine articulations composed of a small, nearly straight vertical ridge proximally bordered by a very small opening encompassed by a slender, low ring; up to four relatively small ambulacral groove spine articulations along ventral plate edge.
Occurrence
Furcaster coulombeae n. sp. is known from the Upper Mississippian (Serpukhovian) Indian Springs Shale Member of the Big Clifty Formation, Crawford County, Indiana, USA.
Description of holotype
Proximal right lateral arm plate CMCIP-94612 (Fig. 6.17–6.19, 6.21, 6.22) higher than long, small, fragile, of triangular outline, with irregularly concave proximal edge lined by a very narrow, poorly defined band of more finely meshed stereom; outer surface composed of coarsely meshed stereom with large, widely spaced pores pointing distalwards at low angle to outer surface, resulting in overall smooth outer surface appearance; distal edge evenly convex and deeply indented by notches of spine articulations, no distal ledge; eight spine articulations deeply sunken in distalwards pointing notches of distal plate edge, with notches equidistant and equally sized and separated by tongue-like distalward somewhat pointed projections of distal edge; spine articulations composed of a small, slender, straight, vertical ridge proximally bordered by a very small opening encompassed by low, slender ring; four moderately large ambulacral groove spine articulations sunken in notches of ventral plate edge; inner side of lateral arm plate with large, conspicuous, strongly prominent and sharply defined vertebral articular knob composed of more finely meshed stereom and projecting dorsalwards from the ventro-proximal plate portion; remaining inner side of lateral arm plate with coarsely reticulate stereom.
Description and variation of paratypes
Median to distal right lateral arm plate CMCIP-94613 (Fig. 6.20, 6.23–6.25) nearly as high as long, of triangular outline, moderately strongly arched; outer surface as in holotype; six lateral and four ambulacral groove spine articulations similar to those of holotype; inner side with vertebral articular knob as in holotype but slightly lower.
Etymology
This species is named in honor of Julie Coulombe who did much of the microfossil picking in support of this project.
Remarks
Furcaster coulombeae n. sp. differs from its congeners in the Sulphur assemblage by having small, thin, fragile, high and strongly arched lateral arm plates with large, widely spaced, distalwards oriented outer surface stereom pores, and up to eight relatively small spine articulations sunken in deep notches of the distal plate edge and composed of a slender, nearly straight vertical ridge and a very small perforation encompassed by a low, thin ring.
Genus Cholaster Worthen and Miller, Reference Worthen and Miller1883
Type species
Cholaster peculiaris Worthen and Miller, Reference Worthen and Miller1883. From the Chesterian of Illinois.
Cholaster sp. indet.
Description
Very large lateral arm plates, proximal examples CMCIP-94550 (Fig. 4.1–4.3) approximately two times higher than long, median to distal examples CMCIP-94551 (Fig. 4.4, 4.5) nearly as long as high; extremely thick and widened (i.e., expanded perpendicular to arm axis), weakly arched, with ventralwards projecting, weakly pointed ventral edge; outer surface with finely meshed stereom devoid of ornamentation; vertical row of spine articulations stretching from the dorsal to the ventral edge and covering almost entire outer surface; spine articulations very large, freestanding, oblique, composed of two prominent, lip-shaped ridges encompassing a slit-like depression; dorsalward increase in size of spine articulations and in distance between them; inner side of lateral arm plates narrow, with a knobby vertebral articular surface ventrally.
Remarks
The lateral arm plates described above stand out because of their enormous size and unusual shape. Strikingly similar lateral arm plates can be seen on specimens from the Mississippian (Chesterian) of Alabama described by Blake and Nestell (Reference Blake and Nestell2019) as Cholaster whitei. Since no other currently known ophiuroid taxon shows even remotely similar lateral arm plates, we assign the material described herein to Cholaster. There are currently two species in the genus, the type species Cholaster peculiaris Worthen and Miller, Reference Worthen and Miller1883, and C. whitei. Differences between the two species (Blake and Nestell, Reference Blake and Nestell2019) are not convincing and pertain to aspects of the dorsal disc skeleton that are easily affected by preservation and/or taphonomic processes. The arm skeleton of the only known C. peculiaris specimen is too poorly preserved to allow for a meaningful comparison with C. whitei. Because of these issues, we prefer to describe our material in open nomenclature, providing an additional record of Cholaster emphasizing that the species-level taxonomy of the genus needs re-assessment.
Both Hunter and McNamara (Reference Hunter and McNamara2017) and Blake and Nestell (Reference Blake and Nestell2019) considered Cholaster as a member of the modern ophiuroid clade, without providing any conclusive comments on the systematic position of the genus. The shape of the lateral arm plates, the spine articulation morphology and the vertebrae with a fully enclosed water vessel canal, however, preclude an assignment to the Ophiovalida, let alone the Metophiurida. Instead, both the lateral arm plates and the vertebrae share similarities with the corresponding ossicles of Onychaster (Hotchkiss and Glass, Reference Hotchkiss and Glass2012). We therefore anticipate close phylogenetic ties between these two genera, pending a more in-depth analysis of their respective morphologies.
Clade Ankhurida Thuy et al., Reference Thuy, Eriksson, Kutscher and Numberger-Thuy2024
Clade Ophiovalida Thuy et al., Reference Thuy, Eriksson, Kutscher and Numberger-Thuy2024
Infraclass Metophiurida Matsumoto, Reference Matsumoto1915
Family unknown
Genus Sulphaster new genus
Type species
Sulphaster odellettorum n. gen. n. sp.
Diagnosis
Same as Sulphaster odellettorum n. sp. by monotypy.
Occurrence
Sulphaster n. gen. is known from the Upper Mississippian (Serpukhovian) Indian Springs Shale Member, Big Clifty Formation, near Sulphur, Crawford County, Indiana, USA.
Etymology
Sulphaster n. gen. named after Sulphur, Indiana, the type locality.
Remarks
Sulphaster n. gen. stands out in the Sulphur ophiuroid assemblage as being very small and fragile. The specimens have a highly characteristic, strongly constricted shape with a bulging proximal edge, a large, thin distal ledge, and a large, deep tentacle notch. No other currently known type of lateral arm plate combines this shape with freestanding spine articulations consisting of a volcano-like cone with a single opening and an irregular striation. We therefore introduce the new genus Sulphaster to accommodate these lateral arm plates.
Sulphaster odellettorum new species

Figure 7. SEM photographs of lateral arm plates; scale bar = 250 μm except (12, 23) = 125 μm; arrows on external views indicate distal direction. (1–12) Sulphaster odellettorum n. gen. n. sp., (1, 2) paratype CMCIP-94571, median LAP, exterior and interior views; (3, 4) paratype CMCIP-94568, distal LAP, exterior and interior views; (5, 6) paratype CMCIP-94569, distal LAP, exterior and interior views; (7, 9–11) holotype CMCIP-94570, proximal left LAP, dorsal, exterior, ventral, and interior views; note the relatively large and well-defined tentacle notch, the large podial basin, the raised spine bases, and the prominent flange on the proximal exterior surface; (8, 12) paratype CMCIP-94567, proximal right LAP, exterior and detail of spine bases views. (13–29) Covidaster medicus n. gen. n. sp. scale bar 500 μm except (28) = 125 μm. (13, 14, 18, 19) holotype CMCIP-94562, proximal LAP, ventral, dorsal, interior, and exterior views; (15–17, 20) paratype CMCIP-94564, median to distal LAP, interior, dorsal, ventral, and exterior views; note the lack of surface ornament; (21, 22, 25, 26) paratype CMCIP-94565, proximal LAP, ventral, dorsal, exterior, and interior views; (23) detail of the ventral pair of spine bases in paratype CMCIP-94566; (24, 27–29) paratype CMCIP-94563, proximal LAP, ventral, dorsal, exterior, and interior views.
Holotype
Holotype specimen CMCIP-94570 (Fig. 7.7, 7.9–7.11) is a proximal left lateral arm plate; Crawford County, Indiana, USA; Indian Springs Shale Member, Big Clifty Formation (Serpukhovian).
Paratypes
Paratypes include CMCIP-94567–94569, 94571; Crawford County, Indiana, USA; Indian Springs Shale Member, Big Clifty Formation (Serpukhovian).
Diagnosis
Small metophiurid with thin, strongly constricted, elongate lateral arm plates showing a bulging outer proximal edge devoid of spurs, a strongly concave dorsal edge, medium-sized, freestanding spine articulations on a slightly bulging distal portion of plate distally bordered by very large, fragile ledge, each spine articulation composed of single, small opening encompassed by a rugose volcano-shaped cone with irregular ridges; dorsalward increase in size of spine articulations; internal side with a very large, deep tentacle notch, and single small, inconspicuous but well-defined, tongue-shaped vertebral articular ridge.
Occurrence
Sulphaster odellettorum n. gen. n. sp. is known from the Upper Mississippian (Serpukhovian) Indian Springs Shale Member, Big Clifty Formation, near Sulphur, Crawford County, Indiana, USA.
Description of holotype
Proximal left lateral arm plate CMCIP-94570 (Fig. 7.7, 7.9–7.11); slightly longer than high, with a strong constriction resulting in a deeply concave dorsal edge and bulging proximal and distal plate portions; distal edge evenly and strongly convex; ventral edge weakly convex to almost straight; outer surface covered by a moderately finely meshed stereom devoid of tubercles; proximal edge evenly concave, lined by a narrow, poorly defined band of more finely meshed stereom, devoid of spurs; four moderately large spine articulations in vertical row, freestanding on bulging distal plate portion, extending from the dorsal to ventral edge and showing dorsalward increase in size of spine articulations and gaps separating them; each spine articulation composed of volcano-shaped cone with single, central opening and irregular, fine striation resulting in an overall rugose aspect; spine articulations distally bordered by large, sharply offset, fragile ledge; internal side of lateral arm plate with small, inconspicuous, well-defined, tongue-shaped vertebral articular ridge at narrowest point between concave dorsal edge and thickened ventral edge; large, deep, well-defined tentacle notch pointing ventralwards and encompassed by thin, poorly defined ridge; inner distal edge slightly pinnacled, devoid of spurs.
Description and variation of paratypes
Median lateral arm plate CMCIP-94571 (Fig. 7.1, 7.2) approximately 1.5 times longer than high, slightly less strongly constricted, otherwise very similar to holotype; three spine articulations similar in shape and position to those of holotype but smaller; internal side as in holotype. Distal lateral arm plate CMCIP-94568 (Fig. 7.3, 7.4) approximately 1.7 times longer than high; similar to holotype except for weaker constriction, much smaller spine articulations and smaller tentacle notch.
Etymology
The species name is derived from the combination of Odell and Odette, the middle names of Archie and Lottie, the nephew and niece of author NSS.
Remarks
The Sulphaster odellettorum n. gen. n. sp. lateral arm plate shape suggesting a complete wrapping of the vertebra, the well-developed, deep tentacle notch and the absence of ambulacral groove spine articulations place it in the clade Ophiovalida comprising all modern-type ophiuroids that lack ambulacral groove spine articulations and have the water vessel canal at least partially open. The shape and position of the spine articulations are strongly similar to Aganaster gregarius (Meek and Worthen, Reference Meek and Worthen1869), implying close phylogenetic ties with Sulphaster n. gen. Therefore, we suggest assignment of Sulphaster odellettorum n. gen. n. sp. to the infraclass Metophiurida, albeit tentatively in the absence of articulated fossils that would allow verifying the clade-diagnostic presence of radial shields and/or genital plates. Considering the shape of the spine articulations (i.e., a single opening encompassed by a volcano-like cone similar to those of Muldaster Thuy et al., Reference Thuy, Eriksson, Kutscher, Lindgren, Numberger-Thuy and Wright2022), we anticipate that Sulphaster n. gen. holds a basal position in the metophiurid stem.
Superorder Ophintegrida O’Hara et al., Reference O’Hara, Hugall, Thuy, Stöhr and Martynov2017
Order Ophiacanthida O’Hara et al., Reference O’Hara, Hugall, Thuy, Stöhr and Martynov2017
Suborder unknown
Family unknown
Genus Covidaster new genus
Type species
Covidaster medicus n. gen. n. sp.
Diagnosis
Same as Covidaster medicus n. sp. by monotypy.
Occurrence
Covidaster n. gen. is known from the Upper Mississippian (Serpukhovian) Indian Springs Shale Member of the Big Clifty Formation, Crawford County, Indiana, USA.
Etymology
Covidaster n. gen. named in memoriam of the Covid-19 pandemic.
Remarks
Covidaster n. gen. lateral arm plates show the spine articulation morphology, consisting of arched, shifted dorsal and ventral lobes encompassing the muscle and nerve openings, that are typically and uniquely found in the members of the extant order Ophiacanthida. The general outline of the lateral arm plates, their outer surface ornamentation, the single spur, and the shape of the vertebral articular ridge on the inner side all favor an assignment to the Ophiacanthida. Similarities are particularly striking with members of the suborder Ophiodermatina, which have more tumid, robust lateral arm plates with small spine articulations. Within the Ophiacanthida, however, relationships are unclear because of the lack of characters that would point to a particular family or allow the erection of a new clade, thus prompting us to leave the suborder and family classification of Covidaster n. gen. open.
Covidaster medicus new species
Holotype
Holotype CMCIP-94562 (Fig. 7.13, 7.14, 7.18, 7.19) specimen is a proximal right lateral arm plate; Crawford County, Indiana, USA; Indian Springs Shale Member, Big Clifty Formation (Serpukhovian).
Paratypes
Paratypes include CMCIP-94563–94566; Crawford County, Indiana, USA; Indian Springs Shale Member, Big Clifty Formation (Serpukhovian).
Diagnosis
Ophiacanthid with small, robust lateral arm plates of rounded outline, weakly arched, relatively low, ventral portion very weakly protruding ventro-proximalwards; outer surface with fine tuberculation; outer proximal edge with single, weak ventro-proximal spur; up to three small spine articulations sunken in shallow notches of distal plate edge and composed of arched, shifted dorsal and ventral lobes encompassing small muscle opening and slightly smaller nerve opening, two ventralmost spine articulations grouped ventrally; internal side with single, tongue-shaped vertebral articular ridge; tentacle opening developed as small, inconspicuous, ventro-distalwards pointing notch in proximal plates, and within-plate perforation in median to distal plates.
Occurrence
Covidaster medicus n. gen. n. sp. is known from the Upper Mississippian (Serpukhovian) Indian Springs Shale Member of the Big Clifty Formation, Crawford County, Indiana, USA.
Description of holotype
Proximal lateral arm plate CMCIP-94562 (Fig. 7.13, 7.14, 7.18, 7.19); plate robust, slightly higher than long, weakly arched, with rounded rectangular outline, with evenly convex dorsal and distal edges; very small ventral portion protruding ventro-proximalwards; proximal edge irregularly concave; outer surface (Fig. 7.19) covered by finely meshed stereom with trabecular intersections merged into very small, uniform tubercles, not showing particular arrangement or pattern; proximal edge lined by narrow, beveled, poorly defined band of slightly more coarsely meshed stereom, devoid of horizontal striation and spurs except for single, prominent spur close to ventro-proximal tip of plate; two equal-sized articulations grouped near ventro-distal tip of lateral arm plate, sunken into distal edge, with third much smaller spine articulation close to dorso-distal tip of plate; spine articulations (Fig. 7.23) composed of arched, shifted dorsal and ventral lobes encompassing a small muscle opening and slightly smaller nerve opening. Internal side of lateral arm plate (Fig. 7.18) with thick dorsal and ventral edges with small contact surfaces with the opposing lateral arm plate, if at all; single, large, prominent, well-defined, tongue-shaped vertebral articular ridge in the center, merged with ventral margin; ventral edge of lateral arm plate distally incised by small, shallow, poorly defined, ventro-distalwards pointing tentacle notch; distal edge of lateral arm plate smooth, pinnacled distally, with single, small, prominent and well-defined spur at ventro-distal tip of plate.
Description and variation of paratype
Median to distal lateral arm plate CMCIP-94564 (Fig. 7.15–7.17, 7.20), as high as long, otherwise similar to holotype in general outline and outer surface ornamentation; three spine articulations as in holotype. Inner side of lateral arm plate as in holotype but with smaller vertebral articular ridge; tentacle opening developed as within-plate perforation.
Etymology
The species named in honor of all members of the medical profession.
Remarks
Covidaster medicus n. gen. n. sp. is by far the oldest representative of the Ophiacanthida, predating the previously oldest member of the clade, Shoshonura brayardi Thuy in Thuy et al., Reference Thuy and Escarguel2019, from the Lower Triassic of Idaho by at least 75 million years, thus pushing the origin of the order into the Carboniferous. This conclusion is consistent with the recent finding that the early diversification of the superorder Ophintegrida, one of the two major living ophiuroid clades, took place much earlier than previously expected (Thuy et al., Reference Thuy, Knox, Numberger-Thuy, Smith and Sumrall2023).
Order Ophioscolecida O’Hara et al., Reference O’Hara, Hugall, Thuy, Stöhr and Martynov2017
Genus Suchaster new genus
Type species
Suchaster granulosus n. gen. n. sp.
Diagnosis
Same as for Suchaster granulosus n. sp. by monotypy.
Occurrence
Suchaster n. gen. is known from the Upper Mississippian (Serpukhovian) Indian Springs Shale Member of the Big Clifty Formation, Crawford County, Indiana, USA.
Etymology
Suchaster n. gen. is a combination of the Greek suchus (Crocodile) and Latin aster (star) referring to the crocodile head-shape of the lateral arm plates.
Remarks
Suchaster n. gen. lateral arm plates are strikingly similar to those recently reported from the Atokan (upper Carboniferous) of Dutton Ranch, Oklahoma (Thuy et al., Reference Thuy, Knox, Numberger-Thuy, Smith and Sumrall2023). They share the same plate outline, outer surface ornamentation, spine articulation morphology, tentacle notch development, and vertebral articular ridge shape, suggesting that they belong to the same genus.
Suchaster granulosus new species

Figure 8. SEM photographs of lateral arm plates; scale bar = 500 μm except (23) = 250 μm; arrows on external views indicate distal direction. (1–23) Suchaster granulosus n. gen. n. sp., (1, 2) paratype CMCIP-94583, distal right LAP, exterior and ventral views; (3, 4) paratype CMCIP-94585, distal right LAP, dorsal and exterior views; (5–7) paratype CMCIP-94584, distal right LAP, interior, ventral, and exterior views; (8, 9, 12, 13) holotype CMCIP-94586, proximal left LAP, ventral, dorsal, exterior, and interior views; note the small, raised spine bases (one dorsal and two ventral) on the distal edge, the well-defined and large tentacle notch, and coarse stereom covering the podial basin and tentacle notch; (10, 11, 14) paratype CMCIP-94588, median right LAP, exterior, ventral, and interior views; (15, 16, 21–23) paratype CMCIP-94587, distal left LAP, dorsal, ventral, interior, exterior, and detail of spine bases views; (17–20) paratype CMCIP-94589, distal right LAP, dorsal, ventral, exterior, and interior views.
Holotype
Holotype specimen is a proximal left lateral arm plate CMCIP-94586 (Fig. 8.8, 8.9, 8.12, 8.13); Crawford County, Indiana, USA; Indian Springs Shale Member, Big Clifty Formation (Serpukhovian).
Paratypes
Paratypes include CMCIP-94583–94585, 94587–94589; Crawford County, Indiana, USA; Indian Springs Shale Member, Big Clifty Formation (Serpukhovian).
Diagnosis
Ophioscolecid with small, elongate, constricted lateral arm plates, higher distally than proximally, with coarsely reticulate outer surface stereom, outer proximal edge with broad band of more finely meshed stereom and two large, prominent spurs; large spine articulations freestanding on slightly bulging distal portion of plate, each composed of single large opening encompassed by horizontal, slightly arched dorsal and ventral lobes; inner side with very large, deep, ventralwards pointing tentacle notch with coarsely reticulate stereom; vertebral articular ridge in proximal third of inner side, relatively small, strongly bent.
Occurrence
Suchaster granulosus n. gen. n. sp. is known from the Upper Mississippian (Serpukhovian) Indian Springs Shale Member of the Big Clifty Formation, Crawford County, Indiana, USA.
Description of holotype
Proximal left lateral arm plate CMCIP-94586 (Fig. 8.8, 8.9, 8.12, 8.13) approximately 1.5 times longer than high, higher distally than proximally, with concave dorsal and ventral edges as result of constriction and bulging distal plate portion, resulting in crocodile-head-like shape; distal edge evenly convex; proximal edge rounded oblique, lined by broad band of more finely meshed stereom and bearing two large, moderately well-defined, elongate, prominent spurs in dorso-proximal and ventro-proximal corners, respectively; outer surface with coarsely reticulate stereom; distal portion of lateral arm plate bulging, with three large, freestanding spine articulations showing dorsalward increase in size and in spacing; spine articulations composed of single, large opening encompassed by large, horizontal, slightly arched dorsal and ventral lobes; spine articulations distally bordered by beveled ledge; dorsal and ventral spine articulations protruding beyond plate outline; internal side with very coarsely reticulate, horizontally stretched stereom in center; relatively small, strongly bent and poorly defined vertebral articular ridge in the proximal one-third of the plate; inner distal edge beveled and bearing two large, moderately well-defined spurs; very large, deep, conspicuous tentacle notch in the middle of ventral edge, pointing ventralwards.
Description and variation of paratypes
Median right lateral arm plate CMCIP-94588 (Fig. 8.10, 8.11, 8.14) approximately 1.8 times longer than high, otherwise similar to holotype in outline and outer surface ornamentation; three spine articulations similar to those of holotype but slightly better preserved, showing smoother dorsal and ventral lobes; inner side as in holotype. Distal left lateral arm plate CMCIP-94587 (Fig. 8.15, 8.16, 8.21–8.23) clearly elongate, two times longer than high, with straight to slightly convex dorsal and ventral edges, otherwise similar to holotype and paratype; inner side as in holotype but with smaller, poorly defined vertebral articular ridge.
Etymology
The species name Suchaster granulosus n. gen. n. sp. refers to the granulose texture of the surface ornamentation.
Remarks
Suchaster granulosus n. gen. n. sp. lateral arm plates described above are strikingly similar to those recently reported from the Atokan (upper Carboniferous) of Dutton Ranch, Oklahoma (Thuy et al., Reference Thuy, Knox, Numberger-Thuy, Smith and Sumrall2023). They share the same plate outline, outer surface ornamentation, spine articulation morphology, tentacle notch development, and vertebral articular ridge shape, suggesting that they belong to the same genus. They differ with respect to the maximum number (three versus four) and arrangement (with a large gap versus continuous) of the spine articulations and the shape of the ventral edge (slightly versus strongly concave), warranting a separation on the species level. We therefore introduce the new genus and species Suchaster granulosus to accommodate the Sulphur material. The lateral arm plates from Dutton Ranch belong to another, yet unnamed species of Suchaster n. gen.
As already fleshed out by Thuy et al. (Reference Thuy, Knox, Numberger-Thuy, Smith and Sumrall2023), Suchaster n. gen. shows a number of synapomorphies of the order Ophioscolecida, representing the oldest member of the order and substantiating the conclusion that the early cladogenesis of the living ophiuroids started as early as the Carboniferous.
Discussion
The Late Paleozoic is considered to be a crucial time in the evolution of Ophiuroidea with the origin of extant clades and demise of the ancestral clades (Hunter and McNamara, Reference Hunter and McNamara2017; Thuy et al., Reference Thuy, Hagdorn and Gale2017). However, the known ophiuroid fossil record is sparse and largely incomplete from the Mississippian to the Triassic (Sprinkle, Reference Sprinkle, Broadhead and Waters1980; Hunter and McNamara, Reference Hunter and McNamara2017; Thuy et al., Reference Thuy, Hagdorn and Gale2017), causing difficulties with timing constraints, modalities, and dynamics of these evolutionary changes. Therefore, a detailed analysis of the intricate morphologies of late Paleozoic ophiuroids is imperative during this evolutionarily complex and important time in the clade’s history.
Our study reveals the presence of 11 ophiuroid species in a single assemblage from the Upper Mississippian (Serpukhovian) of Sulphur, Indiana, USA. This diversity exceeds the previously known global ophiuroid diversity from the entire Mississippian (Spencer and Wright, Reference Spencer, Wright and Moore1966; Sprinkle, Reference Sprinkle, Broadhead and Waters1980, Chen and McNamara, Reference Chen and McNamara2005; Hunter and McNamara, Reference Hunter and McNamara2017). Almost all species are described as new in the present paper, with the exception of one species left in open nomenclature. This highlights the flagrant gap of knowledge on late Paleozoic ophiuroid diversity.
Even more remarkable than the number of species in the Sulphur assemblage is the systematic diversity, comprising one stenuroid (Umerophiura daki n. sp.), two protasterids (Strataster lisae n. sp., Schoenaster limbeckae n. sp.), one cheiropterasterid (Vandelooaster douglasi n. sp.), three furcasterids (Furcaster wardi n. sp., Furcaster mccantae n. sp., and Furcaster coulombeae n. sp.), and three modern-type species (the stem metophiurid Sulphaster odellettorum n. gen. n. sp., the stem ophiacanthid Covidaster medicus n. gen. n. sp., and the stem ophioscolecid Suchaster granulosus n. gen. n. sp.). Furthermore, the presence of three modern-type ophiuroids, both newly described, corroborates that the complex early history of the extant ophiuroid clade started earlier than previously thought (Thuy et al., Reference Thuy, Knox, Numberger-Thuy, Smith and Sumrall2023).
The present study shows that the late Paleozoic ophiuroid diversity is massively underestimated when being assessed based on articulated skeletons alone. Reasons for this major gap between the true ophiuroid diversity on the one hand and the number of species known from intact skeleton fossils on the other mostly revolve around the taphonomic fragility of most taxa and the lack of suitable depositional settings in high-diversity paleoenvironments. We anticipate that most of the species described in the present paper will never be found as articulated skeletons because they occur in settings that fail to facilitate this exceptional type of preservation.
The large knowledge gap of late Paleozoic brittle stars can be remedied by studying the taphonomically less sensitive dissociated ossicles collected from a variety of depositional settings across the late Paleozoic. Our study aligns with the long-established use of dissociated ophiuroid ossicles from Mesozoic and Cenozoic samples (e.g., Jagt, Reference Jagt2000; Thuy, Reference Thuy2013; Thuy and Numberger-Thuy, Reference Thuy and Numberger-Thuy2021) and expands the methods successfully used in these works to the Paleozoic. While the late Paleozoic has considerable amount of work left to be done to better our understanding of brittle star biodiversity, we suggest that it will also be worthwhile to expand these methods into earlier time periods. It is evident that to get a complete picture of brittle star paleobiodiversity, particular attention must be paid to the dissociated skeletal elements throughout the brittle star record.
Acknowledgments
The authors thank University of Tennessee undergraduate J. Coulombe for picking and organizing skeletal material, and graduate student M. Limbeck for help with illustrations. This project was funded by several small grants from The Paleontological Society, Cincinnati Dry Dredgers, Southeastern Section of the Geological Society of America, University of Tennessee Graduate Student Senate, and University of Tennessee Center for Global Engagement. Two reviewers, S. Stöhr and A. Gale, made helpful suggestions that greatly improved this manuscript.
Competing interests
The authors declare none.
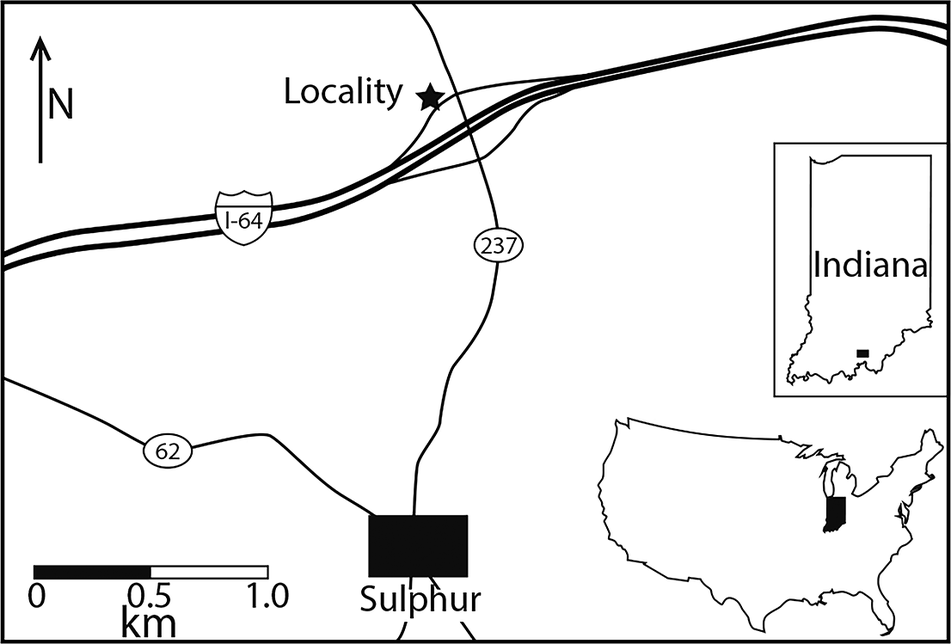




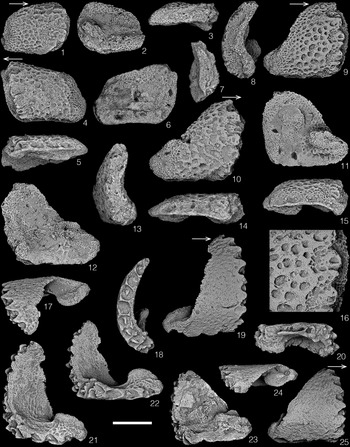



 Open access
Open access