


Non-technical Summary
Macrofossils in Ediacaran sedimentary rocks provide our earliest known evidence of animal morphology. In recent years, much has been learned about the diversity and phylogenetic relationships of Ediacaran metazoans, but many basic aspects of organismic function remain poorly understood. ELEFAN, a statistical package for analyzing length-frequency data, has been shown to provide quantitative insights into the growth dynamics and longevity of living marine invertebrates. Study of a large population of the Ediacaran metazoan Parvancorina minchami, preserved on a single bedding surface exposed in the White Sea region of Russia, permits the inference that despite their unusual morphology and uncertain phylogenetic relationships, these organisms were similar to many living marine invertebrates in terms of their growth and expected longevity of about 4 years. Thus, ELEFAN brings us closer to understanding early metazoans as functioning organisms in Ediacaran ecosystems.
Introduction
Ediacaran macrofossils are simultaneously among the most celebrated and least understood remains known from the fossil record. Although many of these fossils are widely, if not universally, accepted as metazoans (e.g., Narbonne Reference Narbonne2005; Fedonkin et al. Reference Fedonkin, Gehling, Grey, Narbonne and Vickers-Rich2007; Xiao and Laflamme Reference Xiao and Laflamme2009; Sperling and Vinther Reference Sperling and Vinther2010; Erwin et al. Reference Erwin, Laflamme, Tweedt, Sperling, Pisani and Peterson2011; Droser and Gehling Reference Droser and Gehling2015; Dunn et al. Reference Dunn, Liu and Donoghue2018; Hoyal Cuthill and Han Reference Hoyal Cuthill and Han2018; Bobrovskiy et al. Reference Bobrovskiy, Nagovitsyn, Hope, Luzhnaya and Brocks2022; Stephenson et al. Reference Stephenson, Delahooke, Barnes, Rideout, Kenchington, Manica and Mitchell2024), much remains unknown about their anatomy, phylogenetic relationships, and functional biology. Here we employ a method of length-frequency (L/F) analysis known as ELEFAN (Electronic Length-Frequency Analysis) to investigate patterns of growth and mortality in Parvancorina minchami Glaessner, Reference Glaessner1958. ELEFAN was originally developed from first principles to study the growth dynamics of Recent water-breathing ectotherms (WBE; Pauly and David Reference Pauly and David1981), and it was successfully applied to numerous (sub)phyla—here illustrated by one example each—such as fishes (Hikmawansyah et al. Reference Hikmawansyah, Andriani, Khan and Dewanti2019), crustaceans (Dwiponggo et al. Reference Dwiponggo, Hariati, Banon, Palomares and Pauly1986), cephalochordates (Vergara et al. Reference Vergara, Oliva and Riascos2012), echinoderms (Chan et al. Reference Chan, Toh and Huang2018), mollusks (Djan et al. Reference Djan, Fynn and Lazar2022), polychaetes (Omena and Amaral Reference Omena and Amaral2000), cnidarians (Palomares and Pauly Reference Palomares and Pauly2009), and poriferans (Pauly et al. Reference Pauly, Smith and Butler2022). ELEFAN has also been shown to illuminate the growth of extinct marine invertebrates known only through fossils (Pauly and Holmes Reference Pauly and Holmes2022; Wu et al. Reference Wu, Pates, Pauly, Zhang and Fu2024).
The Organisms: Parvancorina minchami
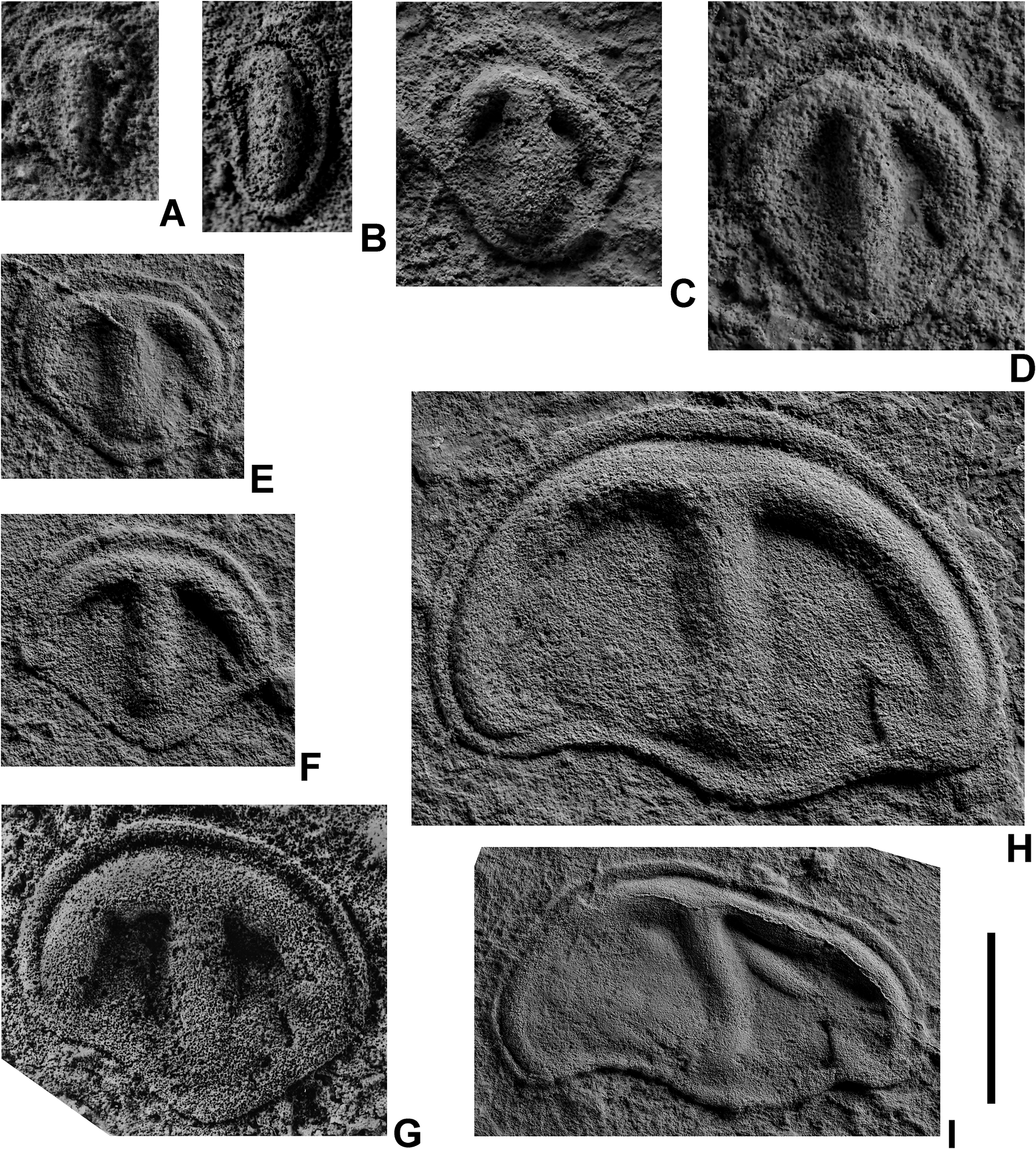
Parvancorina minchami is a distinctive Ediacaran taxon first described by Martin Glaessner (Reference Glaessner1958) from sandstone bedding surfaces of the Pound Quartzite in South Australia. Millimeter- to centimeter-scale individuals have a bilaterally symmetric shield-like morphology, rounded at one end and bluntly pointed at the other. Parvancorina’s most distinctive feature is an anchor-like ridge system, one end curving along the rounded margin and the other running along the central axis of the body (Fig. 1). The only other salient features are fine, parallel lineations of uncertain origin that extend from the axial ridge outward toward the lateral margins (Glaessner Reference Glaessner1980: plates 1,2; Paterson et al. Reference Paterson, Gehling, Droser and Bicknell2017: fig. 4). In addition to Australian occurrences, P. minchami has also been found in Ediacaran sandstones of the Zimnie Gory and Erga Beds along the White Sea coast of Russia (Fig. 2), where it occurs stratigraphically higher than a second, more elongated species, Parvancorina sagitta (Ivantsov et al. Reference Ivantsov, Malakhovskaya and Serezhnikova2004; Naimark and Ivantsov Reference Naimark and Ivantsov2009). The first appearance of Parvancorina in the White Sea section occurs in the underlying Lyamtsa Beds, where it is most likely represented by a new, as yet undescribed species.

Figure 1. Parvancorina minchami Glaessner, Reference Glaessner1958; late Ediacaran (Vendian), southeastern White Sea region, Zimnie Gory locality, burial event horizon Z11(XXII); latex casts from natural imprints with preliminary spraying with ammonium chloride: A, specimen PIN no. 3993/6214; B, specimen PIN no. 3993/6156; C, specimen PIN no. 3993/6286; D, specimen PIN no. 3993/6255; E, specimen PIN no. 3993/9040; F, specimen PIN no. 3993/9021; G, specimen PIN no. 3993/6382; H, specimen PIN no. 3993/8940; I, specimen PIN no. 3993/9019. Scale bar, 2.5 mm (A–D); 0.5 mm (E–H); 10 mm (I).

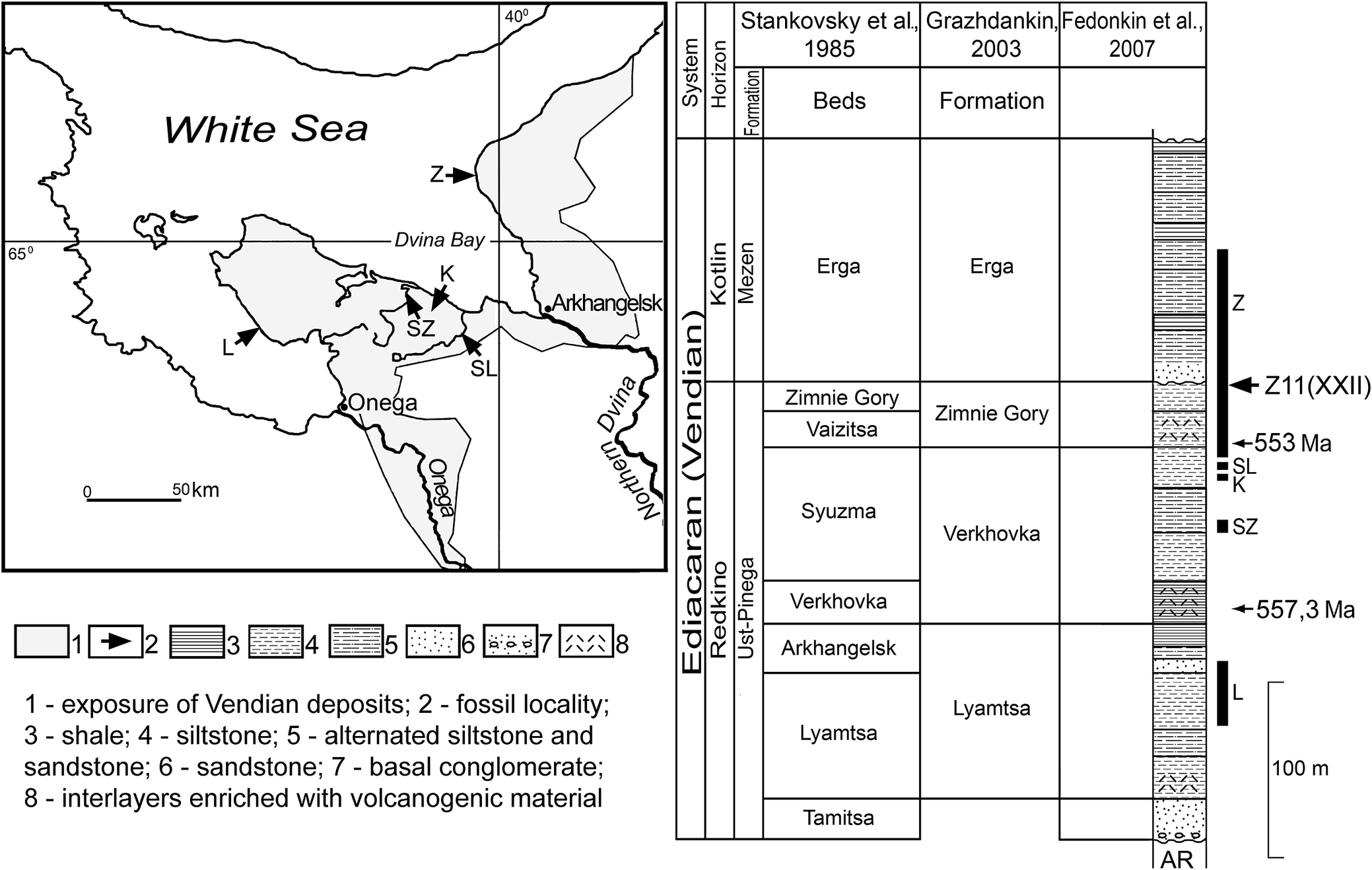
Figure 2. Geographic and stratigraphic position of the burial event horizon Z11(XXII) in the Ediacaran (Vendian) succession of the southeastern White Sea region. Wide vertical stripes to the right of the stratigraphic column correspond to intervals containing macrofossil localities. Letters indicate: L, Lyamtsa; SZ, Syuzma; K, Karakhta; SL, Solza; Z, Zimnie Gory (localities); AR, metamorphic rocks of the Archean and Paleoroterozoic. Stankovsky et al. Reference Stankovsky, Verichev, Dobeiko, Sokolov and Iwanowski1985; Grazhdankin Reference Grazhdankin2003; Fedonkin et al. Reference Fedonkin, Gehling, Grey, Narbonne and Vickers-Rich2007.
Glaessner (Reference Glaessner1958) originally viewed Parvancorina as problematic, but later (Glaessner Reference Glaessner1980) interpreted it as an early arthropod, somewhat distantly related to trilobites. Arthropod affinities were also championed by others, informed by comparisons with marellomorph and other fossils in Cambrian rocks (e.g., Zhang et al. Reference Zhang, Han, Zhang, Liu and Shu2003; Lin et al. Reference Lin, Gon, Gehling, Babcock, Zhao, Zhang, Hu, Yuan, Yu and Peng2006). Careful analysis, however, shows that purported similarities between Parvancorina and arthropods are superficial at best (e.g., Legg Reference Legg2015 and references therein). Like many other Ediacaran macrofossils, Parvancorina shows no evidence of external gills, no mouth or digestive system, and no limbs. Thus, as originally intuited by Glaessner (Reference Glaessner1958), Parvancorina remains problematic, an early macroorganism that exchanged gases by diffusion and likely gained nutrition via phagocytosis by individual epidermal cells, perhaps augmented by osmotrophy.
Several authors have argued that Parvancorina individuals could sense and align themselves with prevailing water movements (Paterson et al. Reference Paterson, Gehling, Droser and Bicknell2017; Coutts et al. Reference Coutts, Bradshaw, García-Bellido and Gehling2018). Employing innovative simulations, Darroch et al. (Reference Darroch, Rahman, Gibson, Racicot and Laflamme2017) made a strong case that such movements would have facilitated the intake of suspended food particles. That said, no trackways document more sustained locomotion, consistent with the absence of limbs or other locomotory structures.
In summary, then, P. minchami was a macroscopic organism capable or orienting itself relative to predominant currents, likely using at least simple muscle fibers. The case is also strong that it gained nutrition by means of suspension feeding. Macroorganisms capable of movement that feed by suspension feeding occur only in the Metazoa, justifying, we believe, the interpretation that P. minchami most likely falls within this kingdom. Even if P. minchami were simply convergent with animals, its structure, habitat, and inferred function would require a physiology not unlike those of unambiguous animals and so amenable to ELEFAN analysis.
Materials and Stratigraphic Setting
This study is based on 211 measured individual Parvancorina minchami out of 651 identified specimens from a single burial event horizon and from a single bedding plane, exposed in outcrops along the White Sea. All specimens are deposited in the Borissiak Paleontological Institute of the Russian Academy of Sciences. The age of this succession is constrained by U-Pb zircon dates for ash layers near the top and bottom of the Zimnie Gory Beds of 550.2 ± 4.6 and 553.0 ± 0.7 Ma, respectively, as well as a date of 557.3 ± 0.6 Ma for the underlying Verkhovka Beds (Martin et al. Reference Martin, Grazhdankin, Bowring, Evans, Fedonkin and Kirschvink2000; Grazhdankin Reference Grazhdankin2016; Yang et al. Reference Yang, Rooney, Condon, Li, Grazhdankin, Bowyer, Hu, Macdonald and Zhu2021). The measured population comes from a single event horizon, Z11(XXII), that preserves an assemblage fossilized in place by instantaneous burial. Such rapid events preserve our closest available approximation to biological populations.
The event horizon Z11(XXII) is found within the Zimnie Gory locality, the northernmost and largest locality among the outcrops of late Ediacaran (Vendian) rocks in the southeastern White Sea region. Stratigraphically, the horizon occurs at the top of the Zimnie Gory Beds of the Ust’-Pinega Formation, or Zimnie Gory Formation in the meaning of Grazhdankin (Reference Grazhdankin2003) (Fig. 2). The burial event horizon can be traced along the southern group of Zimnie Gory outcrops for a distance of at least 2 km. The fossil impressions occur at the base of a lens-shaped layer (0–3.5 cm in thickness) of light gray, fine-grained, cross-bedded sandstone with well-developed ripples on the top. The fossiliferous surface is flat to tuberculate, with a network of thin depressions between the mounds, covered with numerous rounded negative impressions of Aspidella terranovica (which we interpret as attachment structures of the frondose Charniodiscus sp.), with an average diameter of 1–1.5 cm (Fig. 3).

Figure 3. Fossiliferous surface of the burial horizon Z11(XXII), showing an imprint of Dickinsonia costata and five imprints of the attachment disks of Aspidella terranovica. Scale bar, 10 mm.
The Z11(XXII) assemblage is unparalleled for its sheer abundance of benthic Ediacaran biota, including body imprints of Armilifera parva, Charniodiscus sp., Cyanorus singularis, Dickinsonia costata, D. cf. tenuis, Keretsa brutoni, Kimberella quadrata, Cephalonega stepanovi, Ovatoscutum сoncentricum, Paleophragmodictya sp., Paravendia janae, P. minchami, Tamga chamulifera, Tribrachidium heraldicum, Vaveliksia vanaas, and Yorgia waggoneri, as well as enigmatic tubes assigned to Harlaniella ingriana and feeding traces of Dickinsonia, called Epibaion costatus (Ivantsov Reference Ivantsov2013, Reference Ivantsov2017; Zakrevskaya Reference Zakrevskaya2014; Ivantsov and Zakrevskaya Reference Ivantsov and Zakrevskaya2022). Based on many years (1996–2012) of excavations of burial event horizon Z11(XXII), the average concentration of macrofossils is 171 individuals m−2, but in some places, the concentration of Aspidella attachment disks alone reaches 200 individuals m−2 (Zakrevskaya Reference Zakrevskaya2014). Three genera—Kimberella, Dickinsonia, and Parvancorina (e.g., see Ivantsov Reference Ivantsov2009; Naimark and Ivantsov Reference Naimark and Ivantsov2009; Ivantsov and Zakrevskaya Reference Ivantsov and Zakrevskaya2022)—are represented in the collection by several dozens, even hundreds of imprints. Kimberella collected from the burial event horizon Z11(XXII) numbers 770 specimens, ranging from 1.2 to 18.8 mm in width; it had a head section that was rarely preserved. The body of Kimberella was soft, and muscular contraction upon burial led to changes from its generally ribbon-like morphology to an oval or rounded shape. The same is true for Dickinsonia, of which 78 individuals were found, ranging in length from 4 to 145 mm, but which, apparently, also reflect postmortem shape distortion. Thus, the L/F distributions of Kimberella and Dickinsonia cannot be used straightforwardly for reliable inferences about their growth. In contrast, the sharp outline of Parvancorina can be measured accurately and so provides our focus in this study.
The benthic community of the White Sea region lived in and was buried beneath hydrodynamically active shallow waters between fair weather and storm wave bases (Grazhdankin Reference Grazhdankin2004). In general, local burial event horizons extend for several tens, or rarely hundreds, of meters along strike, apparently reflecting the limited volume and trajectory of suspended sediments. In contrast, burial event horizon Z11(XXII) can be traced along strike for 2 km, and we can assume that its area reached several square kilometers. Microbial mat textures are common on bedding plane surfaces (Zakrevskaya Reference Zakrevskaya2014).
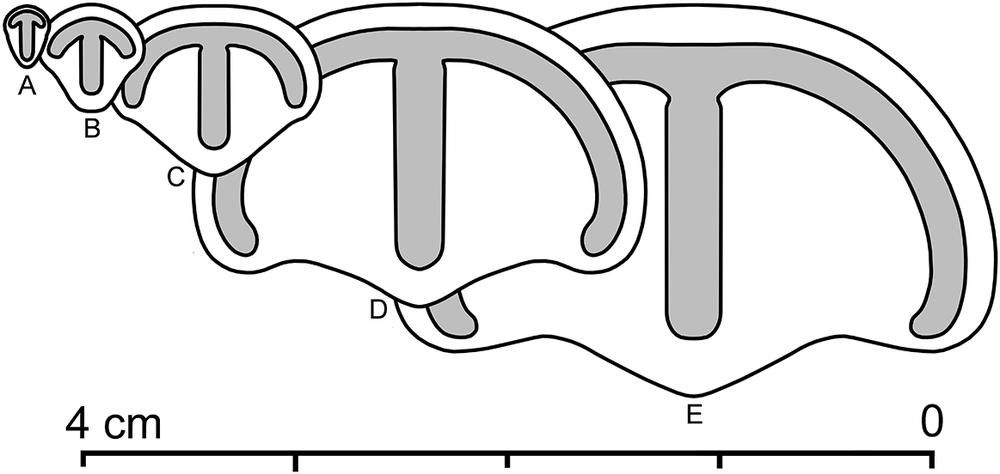
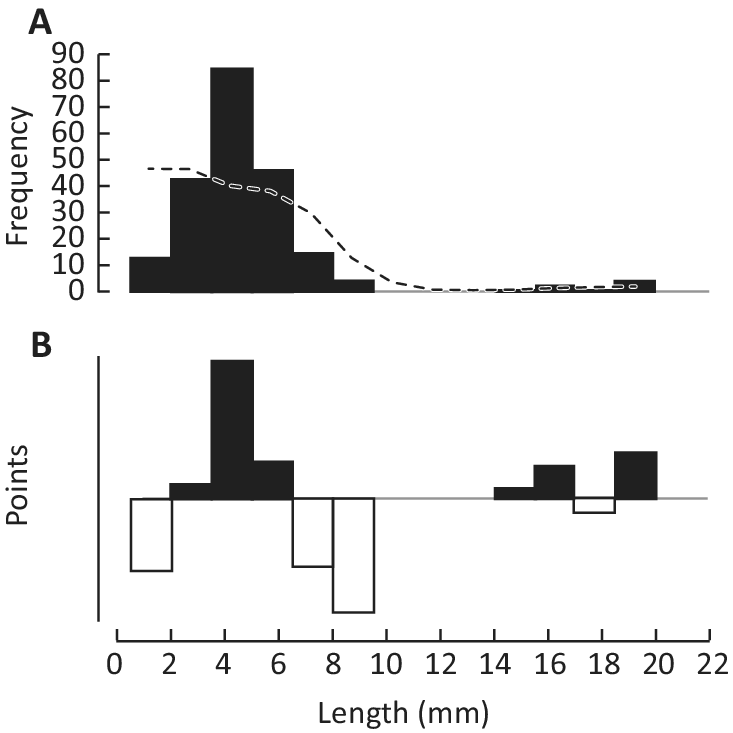
All individuals under consideration were buried in place by a single event, facilitating the collection of reliable data on the growth dynamics and average life span of P. minchami. The lengths and widths of the fossils were measured using plasticine casts of natural imprints using the built-in scale of an MBS-2 binocular microscope, with measured specimens ranging in length from 1.9 to 18.7 mm (Fig. 4). Previously studied populations in White Sea burial horizons commonly demonstrate division into two or more size groups (Zakrevskaya Reference Zakrevskaya2014), presumably reflecting distinct age cohorts and reproductive cycles (Zakrevskaya Reference Zakrevskaya2014; see also Darroch et al. Reference Darroch, Laflamme and Clapham2013; Coutts et al. Reference Coutts, Bradshaw, García-Bellido and Gehling2018). This certainly applies to Parvancorina from the burial horizon Z11(XXII), which fall into two, possibly three, size groupings (Fig. 5A). A clear predominance of small, presumably young individuals may indicate increased mortality of middle-aged and older individuals.

Figure 4. Parvancorina minchami Glaessner, Reference Glaessner1958 from the burial event horizon Z11(XXII); note the change in proportions with increasing size: A, “juvenile” specimen 2.6 mm long, 1.8 mm wide; B, specimen 4.7 mm long, 4.6 mm wide; C, specimen 9.1 mm long, 10.2 mm wide; D, specimen 14.6 mm long, 20.5 mm wide; E, “adult” specimen, 11.8 mm long, 24.6 mm wide. Note allometry, i.e., the width grew faster than the length; see also Naimark and Ivantsov (Reference Ivantsov2009).

Figure 5. Length-frequency (L/F) data for Parvancorina minchami Glaessner, Reference Glaessner1958 from the burial event horizon Z11(XXII), southeastern White Sea region, Russia. A, L/F data of 211 specimens ranging from 2.0 to 18.6 mm, grouped in classes or “bins” of 1.5 mm, with a running average of five classes (dotted line); B, the original L/F data, in each bin, divided by the corresponding running average and 1 subtracted from the quotient. This generates positive (black) points representing peaks in the original L/F data, and negative (white) points, representing the troughs between peaks. Such restructured L/F data are used in ELEFAN (Electronic Length-Frequency Analysis) to estimate the parameters of a best growth curve (see text).
Methods
Ediacaran organisms had no gills or other identifiable dedicated respiratory surfaces. Still, thus, their growth rate (dW/dt) can be conceived as having consisted, as in Recent WBE, of two processes with opposing tendencies, as proposed by Pütter (Reference Pütter1920), yielding
 $$ {d}_W/{d}_t={HW}^d\hbox{--} kW $$
$$ {d}_W/{d}_t={HW}^d\hbox{--} kW $$
where W is the weight (i.e., mass), H is the rate of synthesis of body proteins, d is the exponent of a relationship of the form S ∝Wd, where S is the surface of their bodies exposed to the water that supplied the oxygen required for protein synthesis, and k is the net rate of spontaneous protein denaturation (Pauly and Lam Reference Pauly and Lam2023).
The integration of equation (1) with d = 2/3, that is, assuming that the exposed body surface grew such that S ∝ L 2 while body weight grew according to S ∝ L 3 leads to the von Bertalanffy growth function (VBGF), which is widely used in fisheries and aquatic research (see FishBase, www.fishbase.ca, for fish; SeaLifeBase, www.sealifebase.ca, for marine invertebrates). This has the form:
 $$ {L}_t={L}_{\infty}\left(1-{e}^{-K\left(t-{t}_0\right)}\right) $$
$$ {L}_t={L}_{\infty}\left(1-{e}^{-K\left(t-{t}_0\right)}\right) $$
where Lt is the predicted length at age t, L ∞ is the asymptotic length, that is, the mean length the individuals of a given population would reach if they grew indefinitely, K is the rate, of dimension over time (here: yr−1; with k =3K) at which L ∞ is approached, and t 0 is the hypothetical age at zero length. The parameter t 0 is ignored here, as it is usually very small, and plays no role in the equations that follow.
Note that in Recent WBE, L ∞ is usually slightly larger than L max, the largest individual in a population, and also that longevity (t max) is related to K via the relationship
 $$ {t}_{\mathrm{max}}\approx 3/K $$
$$ {t}_{\mathrm{max}}\approx 3/K $$
Once an estimate of L ∞ is available, K can be estimated by first reconstructing the available length-frequency (L/F) sample using ELEFAN software originally developed by Pauly and David (Reference Pauly and David1981), which multiple applications show is appropriate for WBE reaching up to 10–12 yr, through a procedure involving the following steps:
-
1. selecting a class interval that results in grouping the individual length measurements into 12–15 length classes or “bins” (Fig. 5A);
-
2. calculating the running average of the L/F data over five bins (dotted line in Fig. 5A); and
-
3. dividing the frequency in each bin by the corresponding running average frequency and subtracting 1 from each quotient.
The resulting reconstructed L/F data are shown in Figure 5B, in which the peaks of the original distribution are black positive points, with the troughs originally separating peaks becoming white negative points.
The ELEFAN procedure then selects, from thousands of growth curves generated by a wide range of K values and a single set value of L ∞, the single value of K associated with the growth curve that best links (most of) the peaks while avoiding (as much as possible) the trough of a reconstructed L/F sample, based on the assumption that the sought-after growth pattern repeats itself from one year to the next.
This single best value of K maximizes the number of points that are “hit” by the resulting growth curve. The sum of the positive and negative points that are hit is the explained sum of peaks (ESP), because the curve explains their positions by analogy to the “explained” variance in statistics. The ESP is then related to the available sum of peaks (ASP), that is, the sum of the peaks (i.e., of the highest bins in runs of positive point values, analogous to total variance) via Rn = 10 ESP/ASP )/10, here used as a goodness-of-fit index.
Once an optimal value of K has been estimated, a length-converted catch curve (Pauly et al. Reference Pauly, Moreau and Abad1995) can be used to estimate the instantaneous mortality rate (M) defined by
 $$ {N}_2={N}_1\cdotp {e}^{-M\cdotp {t}_2-{t}_1} $$
$$ {N}_2={N}_1\cdotp {e}^{-M\cdotp {t}_2-{t}_1} $$
and where estimates of M can be obtained from the regression
 $$ \ln \left(N/\varDelta t\right)=a\hbox{--} Mt^{\prime } $$
$$ \ln \left(N/\varDelta t\right)=a\hbox{--} Mt^{\prime } $$
applied to the descending arm of a plot of ln(N/∆t) versus t′ values.
Here, N is the number of specimens in bin i, ∆ is the time needed for the organisms to grow from the lower (L 1i ) to the upper (L 2i ) limit of bin i as estimated for successive values of
 $$ {t}_i=-\left(1/K\right)\cdotp \ln \left[\left({L}_{\infty }-{L}_{2i}\right)/\left({L}_{\infty }-{L}_{1i}\right)\right] $$
$$ {t}_i=-\left(1/K\right)\cdotp \ln \left[\left({L}_{\infty }-{L}_{2i}\right)/\left({L}_{\infty }-{L}_{1i}\right)\right] $$
where ti is the relative age (i.e., the age ignoring t 0) corresponding to successive lengths.
Finally, the age t′ is estimated from
 $$ t^{\prime }=-\ln \left[1-\left({L}_{mr}/{L}_{\infty}\right)\right]/K $$
$$ t^{\prime }=-\ln \left[1-\left({L}_{mr}/{L}_{\infty}\right)\right]/K $$
where L mr is the midrange of a bin.
Table 1 summarizes the parameter definitions and spells out the acronyms used in this contribution. Also note that several versions of ELEFAN versions exist that implement the equations and logic presented above (e.g., Pauly and Greenberg Reference Pauly and Greenberg2013; Taylor and Mildenberger Reference Taylor and Mildenberger2017); the version used here is that of Gayanilo et al. (Reference Gayanilo, Sparre and Pauly2005), available from the Food and Agriculture Organization of the United Nations (www.fao.org/fishery/en/topic/16072/en).
Table 1. Parameters and acronyms used and corresponding definitions.

Results
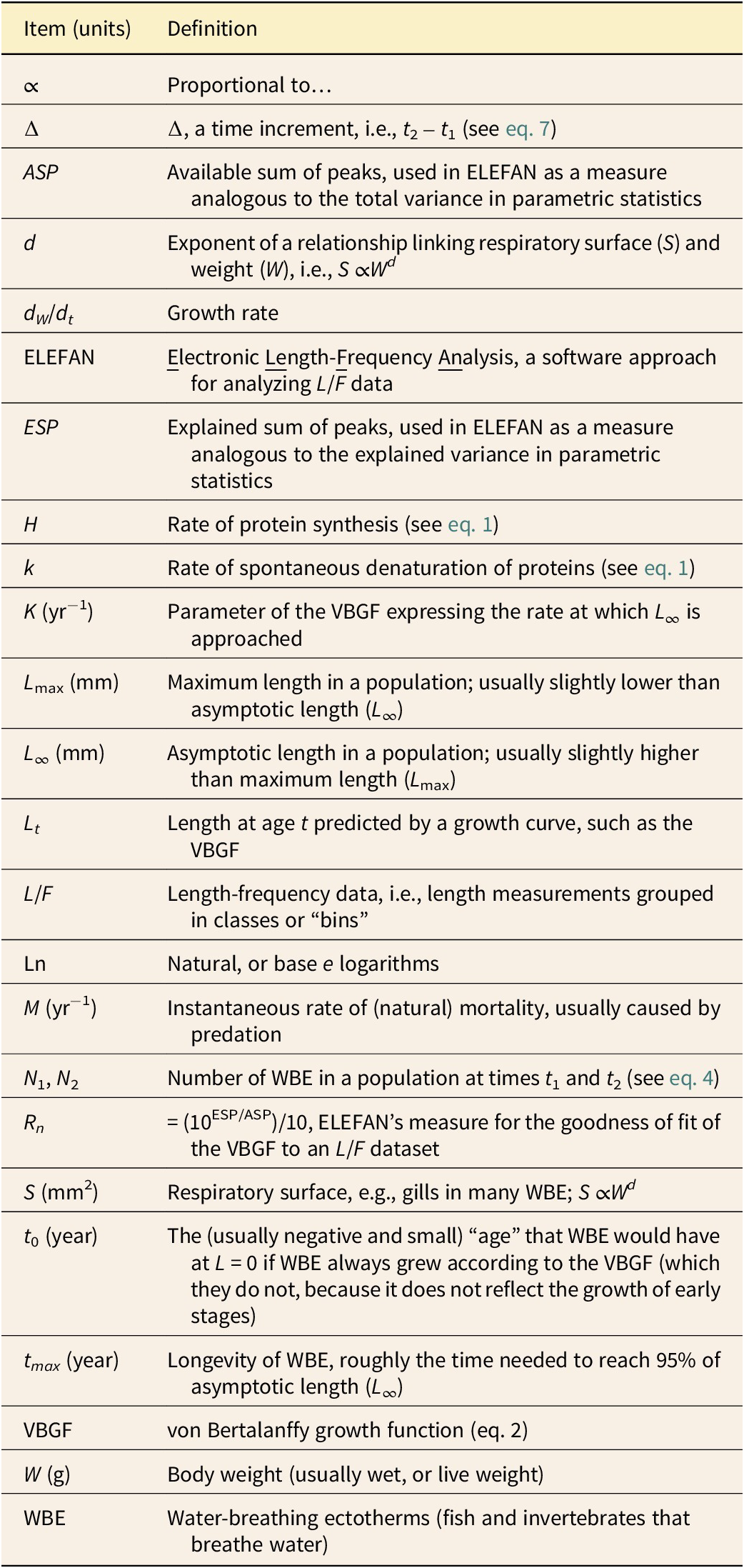
For an asymptotic length set at 20 mm, that is, slightly longer than the maximum reported length of 18.6 mm, and a bin width of 1.5 mm, the K value associated with the highest Rn is K = 0.77 yr−1 (Fig. 6A). This implies, given equation (3), that the longevity of P. minchami was about 4 yr (Fig. 6C). Also, the growth of P. minchami in term of its length was not slower than that of small Recent invertebrates randomly selected as a function of their size from multiple options, such as Thysanoessa longipes (Euphausiacea: Euphausiidae; L ∞ = 17.7 mm, K = 0.72 yr−1), Plesionika martia (Decapoda: Caridae; L ∞ = 24 mm, K = 0.71 yr−1), and Macoma balthica (Mollusca: Bivalvia; L ∞ = 17.1 mm, K = 0.42 yr−1), as can be ascertained from SeaLifeBase (www.sealifebase.ca), based on Iguchi and Ikeda (Reference Iguchi and Ikeda2004), Vafidis et al. (Reference Vafidis, Leontarakis, Dailianis and Kallianiotis2004), and Bachelet (Reference Bachelet1980). The catch curve, constructed with a bin width of 2 mm to generate L/F data that did not contain zeroes), estimated M at 1.14 yr−1 for the specimens larger than 8 mm (Fig. 6B).

Figure 6. Growth and mortality of the Ediacaran Parvancorina minchami Glaessner, Reference Glaessner1958. A, Goodness-of-fit of growth curves featuring asymptotic length (L ∞) of 20 mm, and K-values ranging from 0.1 to 10 yr−1, suggesting that 0.77 yr−1 is the best estimate. B, Catch curve, whose slope for larger “adult” specimens allows the estimation of instantaneous mortality rate M = 1.44 yr−1, which suggests a much higher mortality rate for juveniles. C, Growth curve, as estimated by ELEFAN (Electronic Length-Frequency Analysis) based on the restructured length-frequency data of Fig. 6B, with the Roman numerals referring to various age groups (see text). VBGF, von Bertalanffy growth function.
Discussion
Darroch et al.’s (Reference Darroch, Rahman, Gibson, Racicot and Laflamme2017) elegant study of the relationship of Parvancorina to flowing water was based on digital models of three morphs, P. sagitta, P. minchami from South Australia, and P. minchami from Russia (White Sea). The results indicate that the orientation of these morphs to the direction of the water flow impacted on the turbulence behind their dorsal ridges and thus on the amount of food particles potentially available for deposition and ingestion by phagocytic cells on the rear surface. This simulation therefore strongly suggests that Parvancorina spp. could orient themselves relative to the direction of variable tidal currents and, perhaps, move to places where the water speed and the richness of suspended particles were optimal.
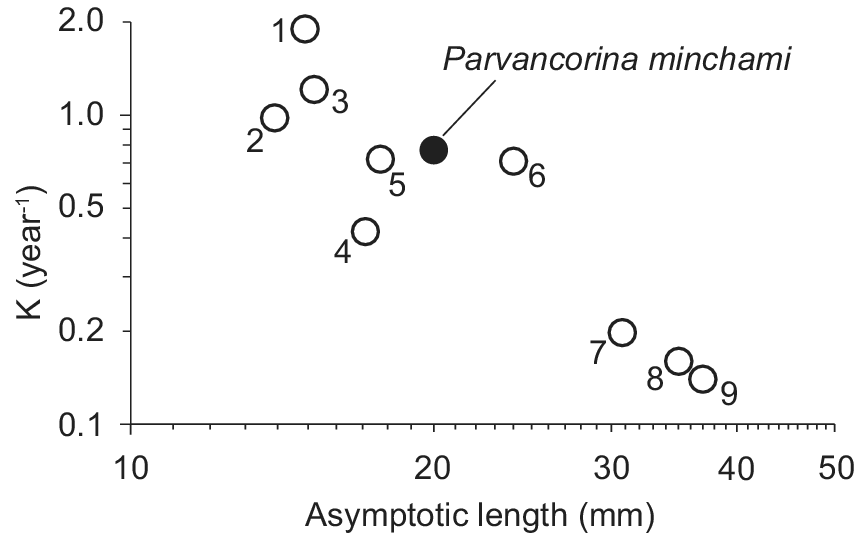
Ineffective as it may seem, this feeding mode, along with the oxygen obtained by diffusion from flowing waters, generated enough metabolic energy for their growth parameters to be comparable to those of small Recent invertebrates (e.g., Apostolidis and Stergiou Reference Apostolidis, Stergiou, Palomares and Pauly2008; see also SeaLifeBase, www.sealifebase.ca), as illustrated in Figure 7. Note, however, that growth, as defined in equation (1), is a process involving mass, and that P. minchami may have been lighter than Recent invertebrates with the same asymptotic length. This, however, would still result in P. minchami having Recent analogues in terms of growth and, hence, metabolic rate. Moreover, these analogue species would also feed on phytoplankton and suspended detritus, the same food that Darroch et al. (Reference Darroch, Rahman, Gibson, Racicot and Laflamme2017) suggested for P. minchami. Possibly, P. minchami was also able to phagocytize organic particles in the underlying sediment on which they rested (M. Laflamme, University of Toronto, personal communication 15 August 2024), which would increase the relative importance of detritivory.

Figure 7. An auximetric plot, allowing the growth parameters of Recent invertebrates whose growth was described by the von Bertalanffy growth function (VBGF; eq. 2) to be compared with Parvancorina minchami. They are: 1, Alpheaus armillatus; 2, Plesionika antigai; 3, Alpheaus estuariensis; 4, Macoma balthica; 5, Thysanoessa longipes; 6, Plesionika martia; 7, Lithophaga patagonica; 8, Arca noae; and 9, Hiatella arctica (see also www.sealifebase.org). This illustrates that P. minchami had a growth performance comparable with that of small Recent invertebrates, at least when their “length” is considered.
The estimated mortality rate of M = 1.14 yr−1 implies M/K ≈ 2, a common ratio in Recent small invertebrates. However, it is clear from Figure 6C that specimens of lengths representing age group II are missing, presumably because of a failure of annual recruitment of young P. minchami to the population represented by the L/F sample (see Houde [Reference Houde, Jakobsen, Fogarty, Megrey and Moksness2016] for possible causes). Thus, our mortality estimates remain tentative. Nonetheless, our estimates dovetail with longevity estimates for a Parvanocorina population from Australia, based on multiple peaks in a size-frequency curve interpreted in terms of seasonal reproduction (Weyland and Droser Reference Weyland and Droser2023).
Conclusions
Length-frequency analysis using the ELEFAN method suggests that Parvancorina minchami had growth and mortality dynamics comparable to those of many small marine invertebrates living today. Thus, despite continuing uncertainties surrounding function and phylogenetic relationships, this brings us a bit closer to an understanding of P. minchami, and by extension other Ediacaran metazoans, as living organisms. More broadly, as demonstrated by previous applications to Paleozoic fossils, the ELEFAN methodology holds the promise of connecting many animal fossils to the huge literature on the dynamics of Recent WBE, enriching our understanding of the paleobiology and ecology of ancient animals and ecosystems.
Acknowledgments
We thank C. Simpson, M. Laflamme, and an anonymous reviewer for helpful comments. We also thank E. Chu for drafting Figs. 5–7. This study was supported by a grant of the Russian Science Foundation (project no. 24-27-00253; https://rscf.ru/project/24-27-00253/) to A.I. and M.A.Z.
Competing Interests
The authors declare no competing interests.


 Open access
Open access