


1. Introduction
Taphonomy (biological, chemical and biostratinomic processes that occur when an organism transfers from the biosphere to the lithosphere) represents a unique and important role within the study of paleobiology (Efremov Reference Efremov1940; Behrensmeyer & Boaz Reference Behrensmeyer, Boaz, Behrensmeyer and Hill1980; Behrensmeyer & Kidwell Reference Behrensmeyer and Kidwell1985; Brandt Reference Brandt1989; Behrensmeyer, Kidwell & Gastaldo 2000; Fernández-Jalvo et al. Reference Fernández-Jalvo, Scott and Andrews2011; Beardmore et al. Reference Beardmore, Orr, Manzocchi and Furrer2012a, Reference Beardmore, Orr, Manzocchi, Furrer and Johnsonb). Konservat lagerstätten, deposits characterized by exceptionally well-preserved, articulated multi-element fossils of skeletons and soft tissues, offer a rare window into taphonomic processes underlying exceptional preservation (Seilacher Reference Seilacher1970; Seilacher et al. Reference Seilacher, Reif and Westphal1985; Bottjer et al. Reference Bottjer, Etter, Hagadorn and Tang2002; Thies & Hauff Reference Thies and Hauff2012; De La Garza et al. Reference De La Garza, Sjövall, Hauff and Lindgren2023; Muscente et al. Reference Muscente, Vinnes, Sinha, Schiffbauer, Maxwell, Schweigert and Martindale2023; Kimmig & Schiffbauer Reference Kimmig and Schiffbauer2024).
One classic example of a konservat lagerstätte that is representative of extraordinary taphonomic phenomena is the Toarcian (Early Jurassic) Posidonienschiefer Formation (= Posidonia Shale), an approximately 183 Ma sedimentary deposit mainly composed of fine-grained black shales that extend across southwestern Germany into France, Luxembourg and Switzerland (Hauff Reference Hauff1921; Birzer Reference Birzer1936; Riegraf et al. Reference Riegraf, Werner and Lörcher1984; Muscente et al. Reference Muscente, Vinnes, Sinha, Schiffbauer, Maxwell, Schweigert and Martindale2023). Hundreds of exceptionally preserved vertebrate fossils have been unearthed from this formation, including fishes, ichthyosaurs, thalattosuchian crocodylomorphs and plesiosaurs (Hauff & Hauff, Reference Hauff and Hauff1981). Overall, thousands of variously preserved fossil vertebrates have been excavated and prepared during the late nineteenth to twentieth centuries, notably in the Holzmaden area of southwestern Germany (Berckhemer Reference Berckhemer1929; Westphal Reference Westphal1961, Reference Westphal1962; Urlichs et al. Reference Urlichs, Wild and Ziegler1994). As such, these fossils represent an important historical palaeontology collection and offer: (1) invaluable archives of past biodiversity on both taxic and community levels; (2) rich sources of raw data and material for a wide range of studies (including palaeobiological, palaeoecological, biogeographical and evolutionary); and (3) document changes in specific palaeocommunities over deep geological time (Lister Reference Lister2011; Vajda & Skovsted Reference Vajda and Skovsted2021). However, detailed information on the preparation of historically collected skeletons is not usually available in museum collections. A notable exception is the collections of the Staatliches Museum für Naturkunde Stuttgart (SMNS), for which some rare information is available, such as how specimens were collected and prepared. This information is crucial, as it can influence specific taphonomic interpretations (e.g. Maxwell et al. Reference Maxwell, Cooper, Mujal, Miedema, Serafini and Schweigert2022), but can also aid in the interpretation of specimens for which no such data are available, as we show in the present study.
Among the vertebrate groups mentioned above, taphonomic analyses have historically focused on ichthyosaurs. This diverse group of Mesozoic marine reptiles with a distinctive thunniform (‘dolphin-shaped’) body plan (e.g. McGowan Reference McGowan1974; Motani Reference Motani2005; Maxwell et al. Reference Maxwell, Fernández and Schoch2012; Reisdorf et al. Reference Reisdorf, Bux, Wyler, Benecke, Klug, Maisch, Fornaro and Wetzel2012; Maxwell & Cortés Reference Maxwell and Cortés2020) are of particular interest in taphonomic studies due to their abundance, unique body form and environmental deposition. Investigations into skeletal articulation and completeness (Hauff Reference Hauff1921; Reisdorf et al. Reference Reisdorf, Bux, Wyler, Benecke, Klug, Maisch, Fornaro and Wetzel2012; Beardmore & Furrer Reference Beardmore and Furrer2016; Beardmore Reference Beardmore2017), soft tissue preservation (Keller Reference Keller1992; Lindgren et al. Reference Lindgren, Sjövall, Thiel, Yheng, Ito, Wakamatsu, Hauff, Kear, Engdahl, Alwmark, Eriksson, Jarenmark, Sachs, Ahlberg, Marone, Kuriyama, Gustafsson, Malmberg, Thomen, Rodríguez-Meizoso, Uvdal, Ojika and Schweitzer2018), seafloor arrival position (Hofmann Reference Hofmann1958; Martill Reference Martill1993; Wahl Reference Wahl2009; Delsett et al. Reference Delsett, Novis, Roberts, Koevoets, Hammer, Druckenmiller and Hurum2016; Maxwell et al. Reference Maxwell, Cooper, Mujal, Miedema, Serafini and Schweigert2022) and the role of their carcasses in benthic enrichment (Martill Reference Martill1987, Reference Martill1993; Danise et al. Reference Danise, Twitchett and Matts2014; Dick Reference Dick2015; Maxwell et al. Reference Maxwell, Cooper, Mujal, Miedema, Serafini and Schweigert2022) present a rich taphonomic exhibition of ichthyosaurs, particularly within the Posidonienschiefer Formation. Specifically, headfirst seafloor arrivals (Hofmann Reference Hofmann1958; Huene Reference Huene1922; Martill Reference Martill1993; Wetzel & Reisdorf Reference Wetzel, Reisdorf, Bromley, Buatois, Mángano, Genise and Melchor2007; Wahl Reference Wahl2009; Delsett et al. Reference Delsett, Novis, Roberts, Koevoets, Hammer, Druckenmiller and Hurum2016; Maxwell et al. Reference Maxwell, Cooper, Mujal, Miedema, Serafini and Schweigert2022), which in the literature are defined as the penetration of the skull at a high angle to the seafloor, present an interesting snapshot of what can occur in the interval between death and burial. In ichthyosaurs, numerous characteristic features have been reported for headfirst arrival specimens (see Discussion below), including dislocation of the skull from the vertebral column, perimortem fracturing in the rostrum and posterior skull, posteriorly displaced dentition and the stratigraphically lower forefin being posteriorly positioned, although this last feature is used with caution (Hofmann Reference Hofmann1958; Wahl Reference Wahl2009; Delsett et al. Reference Delsett, Novis, Roberts, Koevoets, Hammer, Druckenmiller and Hurum2016).
Despite the wealth of taphonomic data on ichthyosaurs from the Posidonienschiefer Formation and other deposits, little is known regarding teleosauroids, another major group of large-bodied vertebrate fossils commonly found within the Holzmaden area. Teleosauroids represent a successful, diverse group of thalattosuchian crocodylomorphs with a specialized set of anatomical features (including a proportionally enlarged skull and femur), range of habitat preferences (including semi-terrestrial, open ocean and freshwater) and unique growth and feeding strategies (Westphal Reference Westphal1961, Reference Westphal1962; Vignaud Reference Vignaud1995; Young & Andrade Reference Young and Andrade2009; Young et al. Reference Young, Hua, Steel, Foffa, Brusatte, Thüring, Mateus, Ruiz-Omeñaca, Havlik, Lepage and Andrade2014; Johnson et al. Reference Johnson, Young, Steel, Foffa, Smith, Hua, Havlik, Howlett and Dyke2017, Reference Johnson, Young and Brusatte2019, Reference Johnson, Young and Brusatte2020, Reference Johnson, Foffa, Young and Brusatte2022, Reference Johnson, Amson and Maxwell2023, Reference Johnson, Scheyer, Canoville and Maxwell2024; Foffa et al. Reference Foffa, Johnson, Young, Steel and Brusatte2019; Martin et al. Reference Martin, Suteethorn, Lauprasert, Tong, Buffetaut, Liard, Salaviale, Deesri, Suteethorn and Claude2019). Teleosauroids were particularly prevalent in the Toarcian, with many ecomorphological factors (such as pseudo-macrophagy, freshwater habitat preference and large body size) established early in their evolutionary history (Westphal Reference Westphal1961, Reference Westphal1962; Li Reference Li1993; Vignaud 1995; Johnson et al. Reference Johnson, Young and Brusatte2020, Reference Johnson, Foffa, Young and Brusatte2022, Reference Johnson, Amson and Maxwell2023, Reference Johnson, Sachs, Young and Abel2025). Numerous specimens of the most common teleosauroid species, Macrospondylus bollensis (Jaeger, Reference Jaeger1828), have been excavated in the Posidonienschiefer Formation (Berckhemer Reference Berckhemer1929; Westphal Reference Westphal1962; Hauff & Hauff Reference Hauff and Hauff1981; Johnson et al. Reference Johnson, Young and Brusatte2020, Reference Johnson, Amson and Maxwell2023, Reference Johnson, Sachs, Young and Abel2025), with body sizes ranging from less than 70 cm to approximately 5 m in length (Johnson et al. Reference Johnson, Amson and Maxwell2023). As such, there is immense potential for taphonomic investigations into teleosauroids from the Posidonienschiefer Formation, which can eventually be compared and integrated with analyses focused on other groups, especially ichthyosaurs. However, aside from brief descriptions of the preservation of individual specimens (e.g. Berckhemer Reference Berckhemer1929; Dridi & Johnson Reference Dridi and Johnson2019; Cortés et al. Reference Cortés, Larsson, Maxwell, Parra Ruge, Patarroyo and Wilson2019), little in-depth taphonomic work has been embarked upon in this group from the Posidonienschiefer Formation or elsewhere. That which has been undertaken is limited to studies on skeletal articulation and completeness (Beardmore et al. Reference Beardmore, Orr, Manzocchi and Furrer2012a), inclusion in a regurgitalite (Serafini et al. Reference Serafini, Gordon, Foffa, Cobianchi and Giusberti2022) and post-burial transportation (Walkden et al. Reference Walkden, Fraser and Muir1987: p. 280).
Here, we provide a comprehensive taphonomic investigation into teleosauroids from the Posidonienschiefer Formation (Fig. 1) with focus on identifying instances of headfirst seafloor arrivals. Throughout the text, we define and refer to ‘headfirst seafloor arrival’ as: individual(s) that display unambiguous skeletal evidence of the head preceding any other anatomical region in penetrating or landing on the seafloor. With the use of ‘unambiguous skeletal evidence’, we refer to any feature in a specimen or 3D model that is associated with a headfirst arrival position and is not contradicted by additional observational data from the same specimen. This definition stems from the results of the present investigation, and it is applicable not only to teleosauroids but also to ichthyosaurs and any other marine vertebrate. We hypothesize that teleosauroid specimens interpreted as headfirst seafloor arrivals display a set of characteristic features associated with this type of landing, and we provide two sets of features indicative of (1) headfirst and (2) non-headfirst seafloor arrivals. We also discuss potential environmental and ecomorphological factors that may have influenced these seafloor landing types and emphasize the importance of understanding previous collection and preparation of historical specimens, which have an effect on environmental and ecological interpretations.
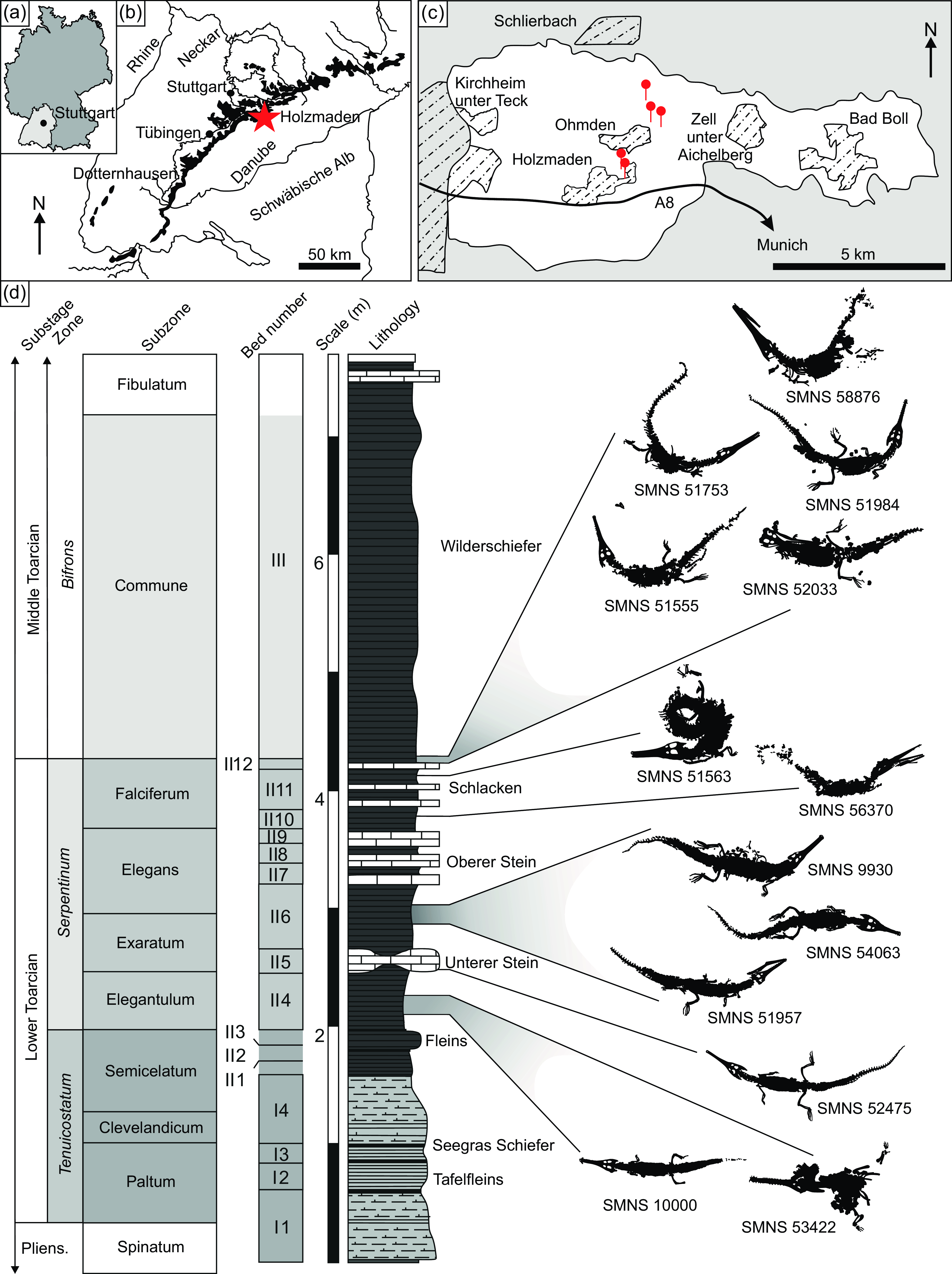
Figure 1. Geological context of teleosauroid specimens from the Posidonienschiefer Formation used in this study. (a) Map of Germany highlighting the state of Baden-Württemberg. (b) Simplified map of Baden-Württemberg showing regional extension of the Posidonienschiefer Formation. The study area of Holzmaden is indicated by a star. (c) Map of the Holzmaden study area with pins indicating the precise collection localities for specimens whereby this data is recorded. (d) Stratigraphic log of the Posidonienschiefer Formation based on the section at Holzmaden and correlated to ammonite biozones. The collection horizon of each specimen used in this study is represented with silhouettes. Maps and logs are modified from Riegraf et al. (Reference Riegraf, Werner and Lörcher1984), Maxwell et al. (Reference Maxwell, Cooper, Mujal, Miedema, Serafini and Schweigert2022) and Cooper et al. (Reference Cooper, Giles, Young and Maxwell2022).
2. Geological setting
The Lower Jurassic (lower–middle Toarcian) Posidonienschiefer Formation crops out extensively in Germany between Lower Saxony in the north, to Waldshut-Tiengen in the south, with additional exposures of coeval marine sedimentary deposits in Austria, Luxembourg, France, the UK and Switzerland (Howarth, Reference Howarth1962; Riegraf et al. Reference Riegraf, Werner and Lörcher1984; Röhl and Schmid-Röhl Reference Röhl and Schmid-Röhl2005; Bour et al. Reference Bour, Mattiolo and Pittet2007; Trabucho-Alexandre et al. Reference Trabucho-Alexandre, Dirks, Veld, Klaver and De Boer2012; Williams et al. Reference Williams, Benton and Ross2015; Mönnig et al. Reference Mönnig, Fraz and Schweigert2018). All of the specimens featured in this study originate from the area of Holzmaden in Baden-Württemberg, southern Germany (approximately 10 km SE of Stuttgart) (Fig. 1a–b). The label of ‘Holzmaden’ is often used colloquially to refer to any number of active or former quarries in or around the villages of Kirchheim unter Teck, Jesingen, Holzmaden, Ohmden, Schilerbach, Zell unter Aichelberg and Bad Boll (Fig. 1c) (Maxwell et al. Reference Maxwell, Cooper, Mujal, Miedema, Serafini and Schweigert2022; Cooper et al. Reference Cooper, López-Arbarello and Maxwell2024). The stratigraphic section at Holzmaden represents a continuous succession of bituminous mudstones, compact oil shales and marly limestones spanning between the Tenuicostatum – Bifrons ammonite Zones of the early to middle Toarcian (e.g. Riegraf et al. Reference Riegraf, Werner and Lörcher1984). The section records the onset and fallout of the Jenkyns Event; an apparent small-scale extinction event marked by a temporary global depletion in seafloor oxygenation and extensive deposition of pyrite-rich black shales in the Serpentinum Zone. Thalattosuchian crocodylomorphs are rare in the formation but are continuously distributed between beds ϵII3 (‘Fleins’) and ϵII12 (‘Schlacken’), corresponding to a temporal range between the uppermost Tenuicostatum Zone (Semicelatum Subzone) and the uppermost Serpentinum Zone (Falciferum Subzone) (Hauff Reference Hauff1921). All fourteen specimens featured in this study were collected from quarries in the vicinity of Holzmaden and Ohmden (Fig. 1c) from various stratigraphic horizons in the Serpentinum Zone (beds ԑII4 – ԑII12; Fig. 1d).
3. Materials and methods
3.a. Institutional abbreviations
FMNH, Field Museum of Natural History, Chicago, USA
GPIT, Paläontologische Sammlung der Eberhard Karls Universität, Tübingen, Germany
GZG, Geologisches Institut Geologisch-Paläontologisches, Göttingen, Germany
MMG, Staatliches Museum für Mineralogie und Geologie, Dresden, Germany
NMWIN, Naturmuseum Winterthur, Winterthur, Switzerland
NHMW, Naturhistorisches Museum Wien, Vienna, Austria
OUMNH, Oxford University Museum of Natural History, Oxford, UK
PIN, Paleontological Institute, Moscow, Russia
PMU, Evolutionsmuseet Uppsala Universitet, Uppsala, Sweden
RNHM F 6254, Naturkundemuseum Reutlingen, Reutlingen, Germany
SMNS, Staatliches Museum für Naturkunde Stuttgart, Stuttgart, Germany.
UMH, Urweltmuseum Hauff, Holzmaden, Germany
3.b. Skeletal material
Fourteen teleosauroid specimens (thirteen representing the basal machimosaurid Macrospondylus bollensis and one representing the teleosaurid Platysuchus multiscrobiculatus) from the Posidonienschiefer Formation and housed in the collections of the Staatliches Museum für Naturkunde Stuttgart (SMNS) were analysed first-hand to assess and identify individuals exhibiting characters associated with headfirst seafloor arrival (Supplementary Table 1). These specimens were subjected to photogrammetry to enhance the observation of features (see below; Supplementary Table 1). In addition, we examined a total of 28 specimens at the SMNS and additional collections, either first-hand or through provided photographs, to review the frequency of occurrence of headfirst seafloor arrival in thalattosuchians, while also testing the methods here employed to differentiate arrival positions.
3.c. Photogrammetry
Three-dimensional (3D) photogrammetric models were created for the fourteen teleosauroid specimens studied in detail (Supplementary Table 1). This technique creates 3D surface models of a given object, by taking photographs (1) from all perspectives and/or (2) perpendicular to the surface with consecutive photographs overlapping each other by approximately two-thirds. Detailed explanations on photogrammetry and specifically on how to obtain photographs are provided by Mallison & Wings (Reference Mallison and Wings2014) and Lallensack et al. (Reference Lallensack, Buchwitz and Romilio2022). Here, we used digital cameras Canon EOS 1300D (device standard lens EFS 18–55 mm) and Canon PowerShot SX410 IS (device standard lens 4.3–172 mm F.3.5–F6.3). The number of photographs taken for each specimen (Supplementary Table 1) varies according to size and complexity and details needed from the fossil.
Three software packages were used to produce the 3D models and their final rendering, mainly following the workflow by Mujal et al. (Reference Mujal, Marchetti, Schoch and Fortuny2020) and Maxwell et al. (Reference Maxwell, Cooper, Mujal, Miedema, Serafini and Schweigert2022): (1) Agisoft Metashape v.1.8.3 (professional license, educational version; https://www.agisoft.com/), to create the dense cloud, mesh and texture; (2) MeshLab v.2020.07 (open access; https://www.meshlab.net/) to scale, clean and orientate the mesh and (3) ParaView v.4.1.0 (open access; https://www.paraview.org/), to produce false-colour depth maps and contours.
To obtain information related to the orientation of each individual when landing on the original substrate, 3D models were oriented (in MeshLab) according to the original stratigraphic plane by identifying layers and/or laminae distinguishable within the sedimentary matrix (Fig. 2). First, stratigraphic planes were identified for each specimen by examining the original surfaces of the strata or laminae containing each specimen; the same planes were then also located in the 3D models. Then, the automatic orientation in MeshLab was used, aligning each model to the XY plane (thus, with the z-axis perpendicular to the model). In most cases, the automatic alignment (plus rotation) to the XY plane in MeshLab coincides with the sedimentary plane. In those models not correctly aligned automatically, a manual re-alignment to the stratigraphic plane was done also in MeshLab. In these cases, a rotation of only 0.5º to 1º to the x-axis, y-axis or both was required. Afterwards, the orientation of each model was checked in ParaView by applying contours on the model and comparing them with the textured model. If any corrections were necessary, 3D models were oriented again in MeshLab until obtaining an accurate result.

Figure 2. Orthophotographs of the 3D photogrammetric model of SMNS 53422. (a) Image with original texture. (b) Image with original texture and contours. Arrows in the close ups (a’ and b’) point to distinguishable sedimentary layers that coincide with the contours, showing that the model is aligned with the XY plane.
Finally, prepared 3D models were rendered in ParaView to produce images in orthogonal view with different filters: (1) original texture, to compare with and track details from the original fossil; (2) false-colour depth map (aligned to the z-axis), to determine the relative stratigraphic position of elements and (3) contours (aligned to the z-axis, and plotted on the original texture and on the depth map) to enhance orientation and relative stratigraphic position of elements. For the false-colour depth maps, different preset colour scales from ParaView v.4.1.0 were used: ‘Rainbow Desaturated’, ‘gist_earth’ and ‘Cool to Warm’, with their respective colour spaces being RGB, CIELAB and Diverging. Figures in the present work include false-colour depth maps in ‘Rainbow Desaturated’. The other two preset colour scales can be distinguished by readers with colour vision deficiency (see Lautenschlager Reference Lautenschlager2021); in the supplementary material (Supplementary Figs. 1–5) all the images of the 3D models with ‘gist_earth’ colour scale are provided. The number of contours used varies in each specimen according to the most appropriate visualization, usually ranging from 25 to 60 lines.
3.d. Taphonomic analysis
The disposition of fractured bones was examined to determine the origin of the damage (diagenetic vs. perimortem vs. postmortem; e.g. Hofmann Reference Hofmann1958; Green & Schultz Reference Green and Schultz2017; Spiekman & Mujal Reference Spiekman and Mujal2023) and test them against the seafloor landing posture of each individual. Peri- and postmortem fractures show physical displacement of the broken fragments, with sediment accumulated within the break (Hofmann Reference Hofmann1958). In contrast, diagenetic fractures show compaction-related breakage without substantial displacement of the fragments, and with only diagenetic mineral deposits – if any foreign material at all – within the fracture. Peri- and postmortem fractures in bone can be differentiated by the geometry of the broken ends of the bone (e.g. Green & Schultz Reference Green and Schultz2017; Spiekman & Mujal Reference Spiekman and Mujal2023). Geometry and orientation of the fractures with respect to the bones as well as the general posture of each specimen were taken into account. In addition, the relative stratigraphic position and orientation of elements (with respect to both the substrate and the rest of the skeleton), stratigraphic horizon and lithology were assessed. A relevant criterion to interpret the stratigraphic orientation is the articulation and relative position of skeletal elements: the stratigraphically lower skeletal elements are usually articulated (e.g. osteoderms, ribs and limbs from one side in individuals lying laterally on the substrate), because those were not exposed to the environment, and thus, decay was minor or slower than in the exposed parts of the skeleton. Moreover, the sediment would have served a stabilizing function, preventing these elements from being displaced even in the presence of decay. Therefore, in specimens prepared from the topside, the articulated elements are the lowest ones in the slabs (i.e. still embedded in the sedimentary matrix; e.g. SMNS 51555), whereas in specimens prepared from the underside it is the opposite (i.e. mostly free of the matrix; e.g. SMNS 9930).
4. Results and interpretations
Specimens examined mainly include complete to partially complete skeletons that are well articulated or with skeletal elements still in anatomical position. This is because features to determine whether an individual represents a headfirst seafloor arrival or not are best observed on specimens that remained mostly articulated until complete burial. In this sense, massive skeletal disarticulation of a specimen can make it difficult to distinguish between perimortem headfirst seafloor arrival features (particularly those involving displacement of cranial and vertebral elements), and general postmortem taphonomy (e.g. SMNS 15816). Therefore, specimens that are (1) highly disarticulated or (2) display a high degree of disorganized or random orientation of the skeletal elements due to the normal or usual decay processes of a vertebrate carcass (Beardmore et al. Reference Beardmore, Orr, Manzocchi and Furrer2012a; Beardmore & Furrer Reference Beardmore and Furrer2016; Orr et al. Reference Orr, Adler, Beardmore, Furrer, McNamara, Peñalver-Mollá and Redelstorff2016) are not considered here, since some preservational features might be misinterpreted and/or obliterated. It is also important to note that the penetration angle of the animal into the substrate is inconsequential; instead, the notion that the head did not impact the substrate horizontally but rather at an angle, prompting fracturing of the skull and penetration into the substrate is emphasized.
One of the fourteen specimens examined in detail corresponds to a juvenile individual of Macrospondylus bollensis (SMNS 15391) that, although being a nearly complete skeleton, cannot be determined as headfirst or non-headfirst seafloor arrival because of the way it was (historically) prepared. This issue is further explained in the Discussion below.
4.a. Headfirst seafloor arrivals
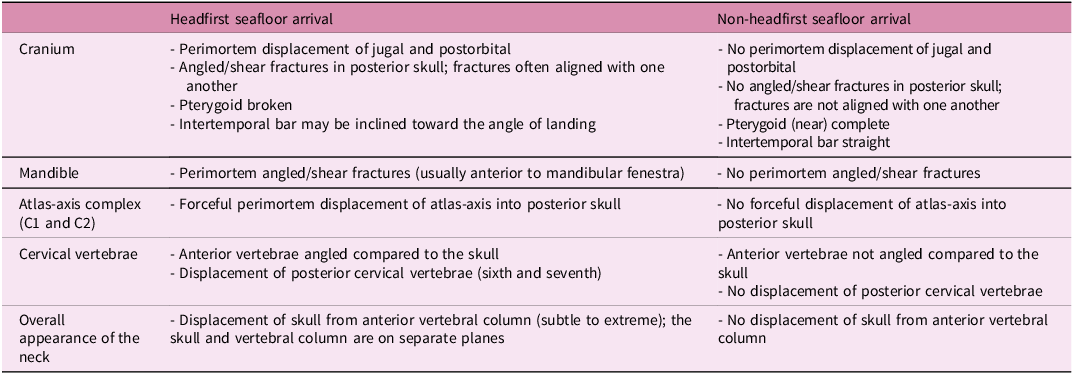
Out of thirteen 3D-modelled SMNS specimens, ten are interpreted as headfirst seafloor arrivals as defined in the Introduction (see above). There are numerous unambiguous features that we associate with a headfirst seafloor arrival (Figs. 3, 4; Table 1; Supplementary Figs. 1–4). These features are attributed to either (a) force applied parallel to the long axis of the skull, as in an anterior collision with the seafloor, or (b) unequal taphonomic deformation related to the skull becoming embedded at an angle relative to the bedding plane. Of note, not all features are present (or visible) in all specimens, which may be related to different biological (e.g. ontogeny) and/or environmental factors (see Discussion below). Regardless, the presence of at least one of these features (if not otherwise indicated) is sufficient to identify the specimen as a headfirst seafloor arrival individual. These distinct features include:
-
1. Disarticulation and anterior or posterior displacement of the cranium and mandible from the cervical region (= neck) of the vertebral column (Fig. 3a–d). The skull (= cranium + mandible) is situated on a different plane than the anterior vertebral column (i.e. the skull and neck diverge in different directions). This displacement can be either extreme (Fig. 3a, b) or subtle (Fig. 3c, d). Importantly, the observation and interpretation of this feature also depends on the orientation of the specimen with respect to the substrate (e.g. if the skull is oblique to the substrate, as in SMNS 51753, the displacement is less evident – or subtle – due to diagenetic compression and compaction).
-
2. Displacement and fracturing of small or thin cranial bones such as the lacrimal, prefrontal, jugal, postorbital and/or quadratojugal bar (Fig. 3e). In addition, slight dislocation immediately anterior or posterior to the orbits may occur. For example, the posterolateral rim of the left orbit in SMNS 51555 has been slightly shifted anteriorly, causing the left orbit to appear smaller than that on the right (Fig. 3f). The displacement or shifting of these skeletal elements is correlated with the direction and/or orientation in which the individual landed on the seafloor (see Table 2).
-
3. Angled, perimortem fractures in the posterior portion of the cranium (= posterior to the orbits), most notably in the posterolateral border of the supratemporal fenestrae. Often the fractures align with one another along the same plane (Fig. 3g), also indicating the direction in which the individual landed on the seafloor.
-
4. Slight inclination of the intertemporal bar toward the interpreted landing side may be present (Fig. 3d). This feature on its own is insufficient to infer landing posture but is often associated with more diagnostic characters indicative of headfirst seafloor arrival.
-
5. When visible, the pterygoid usually shows substantially perimortem breakage and displacement (Fig. 3e). This feature on its own is insufficient to infer landing posture but is often associated with more diagnostic characters indicative of headfirst seafloor arrival.
-
6. Angled perimortem fractures in the mandible, often immediately anterior and posterior to the mandibular fenestra (Fig. 3g). As in the cranium, the fractures often align with one another along the same plane, indicating the direction in which the individual landed on the seafloor.
-
7. In extreme cases, the two rostral and/or hemimandibles may be separated (Fig. 4a). However, this feature is treated with caution, as many taphonomic factors may contribute to this disassociation (such as carcass decay prior to final burial or extent of ossification due to biological [= ontogenetic] stage [i.e. sutures are less tightly closed in juveniles]).
-
8. Forceful perimortem displacement of the atlas-axis complex (cervical vertebrae one and two) into the posterior part of the skull (Fig. 3g). In extreme cases, this can result in the posterior skull becoming obliterated (Fig. 3e).
-
9. Perimortem displacement of the last two cervical vertebrae (= 6th and 7th) from the atlas-axis complex (= the 1st and 2nd cervical vertebrae) and remaining cervical vertebrae (Fig. 4b). This feature is usually correlated with either a curving or ‘L-shaped’ appearance to the entire neck (Figs. 3a–d, 4a, b).

Figure 3. Features of headfirst seafloor arrival teleosauroids 1. (a–d) Orthophotos (a, c) and false-colour depth maps (‘Rainbow desaturated’) (b, d) of the skull and neck area from the 3D models of SMNS 51563 (A, B) and SMNS 51753 (c, d). White lines in (a) and (c) represent disarticulation and displacement of skull (solid line) and neck (dashed line) resulting in diverging orientations. In (d), the intertemporal bar is inclined toward the landing side, which is indicated by the inclination of the skull. (e) Posterior part of the skull and anterior part of the neck of SMNS 56370 showing fractured elements and displacement of the vertebrae into the skull. (f) Posterior part of the skull of SMNS 51555 showing orbits with different sizes (left orbit smaller than the right one, highlighted with solid and dashed lines) and a fracture of right temporal bar (arrowed). (g) Posterior part of the skull of SMNS 51957 showing multiple fractures that are aligned (arrowed and marked with dashed lines). (h) Close up of the posterior part of the skull of SMNS 53422 showing multiple fractures on the mandible that are aligned (arrowed and marked with dashed lines). Note that SMNS 51563 was originally modelled by H. Mallison and recently modified by E.M.

Figure 4. Features of headfirst seafloor arrival teleosauroids 2. (a) Orthophoto of the skull from the 3D model of SMNS 52033; note that the mandible is split through the symphysis. (b) Orthophoto of the skull and neck from the 3D model of SMNS 9930; note that vertebrae are displaced giving an ‘L-shaped’ appearance to the neck (marked with dashed line), with a clear breakage between 7th and 8th vertebrae (arrowed). (c, d) Orthophotos of the 3D model of SMNS 10000 with original texture (c) and false-colour depth map (‘Rainbow desaturated’) (d). In D, the inclination of the skull (oblique to the substrate) is well visible, encompassing most of the range of colours of the depth scale bar (indicated with the elongated triangle and plus and minus symbols). Note that SMNS 9930 was originally modelled by H. Mallison and recently modified by E.M.
Table 1. Headfirst vs. non-headfirst seafloor arrival features in Posidonienschiefer Formation teleosauroids
Regarding the orientation of the specimen with respect to the bedding plane, there are multiple interpreted carcass embedding postures associated with headfirst seafloor arrival (Table 2). These include: (1) dorsolateral landing (a) right (e.g. SMNS 9930) or (b) left (e.g. SMNS 51563); (2) ventrolateral landing (a) right (e.g. SMNS 51555) or (b) left (e.g. SMNS 56370); and (3) general ventral landing (e.g. SMNS 52033). In addition, the elevation maps obtained from the 3D models show that, in some cases, the skull lies at an angle, with the tip of the snout being stratigraphically lower than the back of the skull (Figs. 3d, 4c, d), a feature commonly observed in headfirst seafloor arrival of ichthyosaurs (e.g. Martill Reference Martill1993, Reference Martill1987; Maxwell et al. Reference Maxwell, Cooper, Mujal, Miedema, Serafini and Schweigert2022). It is important to note that there is no radical displacement or deformation of the dentition, in contrast with what is observed in some ichthyosaurs (see Discussion below). This is attributed to clade-specific differences in tooth implantation.
Table 2. Interpreted seafloor arrival type, seafloor landing and preparation in Posidonienschiefer Formation teleosauroids
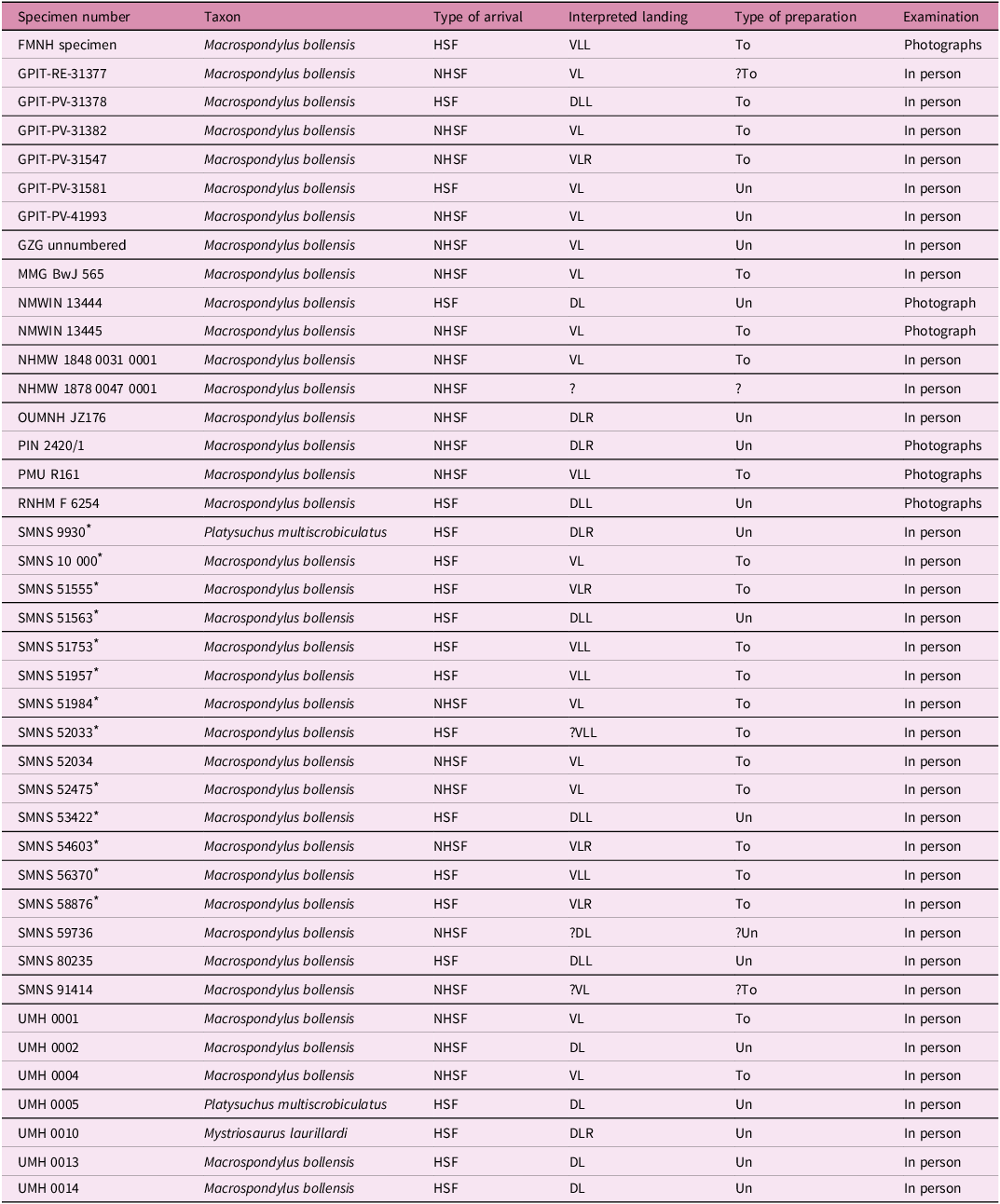
Abbreviations: DL = general dorsal landing; DLL = dorsolateral left landing; DLR = dorsolateral right landing; HSF = headfirst seafloor arrival; NHSF = non-headfirst seafloor arrival; To = topside preparation; Un = underside preparation; VL = general ventral landing; VLL = ventrolateral left landing; and VLR = ventrolateral right landing. Note that non-headfirst seafloor arrivals do not contain unambiguous features seen in headfirst seafloor arrivals (Table 1); however, this does not mean that the head was not the initial point of contact with the seafloor (see text for discussion). An asterisk (*) indicates that the specimen was used for a 3D photogrammetric model.
As indicated above, to assess that a specimen represents a headfirst seafloor arrival, it is not necessary to identify all these features. For example, the juvenile specimen SMNS 10000 only displays the head in a stratigraphically lower position with respect to the postcranium, being also obliquely oriented with respect to the stratigraphic surface (Fig. 4c, d).
4.b. Non-headfirst seafloor arrivals
Out of thirteen 3D-modelled SMNS specimens, only three are interpreted as non-headfirst seafloor arrivals (Fig. 5), all presenting nearly the same set of visibly distinct features that we associate with a non-headfirst seafloor arrival (Table 1). These include:
-
1. Lack of displacement of the cranium from the anterior cervical region. Both the cranium and mandible are on the same plane as the anterior vertebral column.
-
2. Little to no displacement of cranial bones, which are preserved in situ. Note that this does not apply to specimens that underwent a relatively prolonged decay process; in those specimens, determining whether they are headfirst seafloor arrivals or not is usually not possible.
-
3. Absence of noticeable angled perimortem fractures in the posterior cranium.
-
4. No intertemporal bar inclination.
-
5. When visible, the pterygoid is generally well-preserved and in situ (Fig. 5a).
-
6. Lack of angled perimortem fractures in the mandible. The bones surrounding the mandibular fenestra (dentary, angular and surangular) are generally intact.
-
7. The rostral and mandibular halves are sutured together.
-
8. Absence of atlas-axis (1st and 2nd cervical vertebrae) displacement into the posterior skull.
-
9. The last two cervical vertebrae are on the same plane as the rest of cervicals.
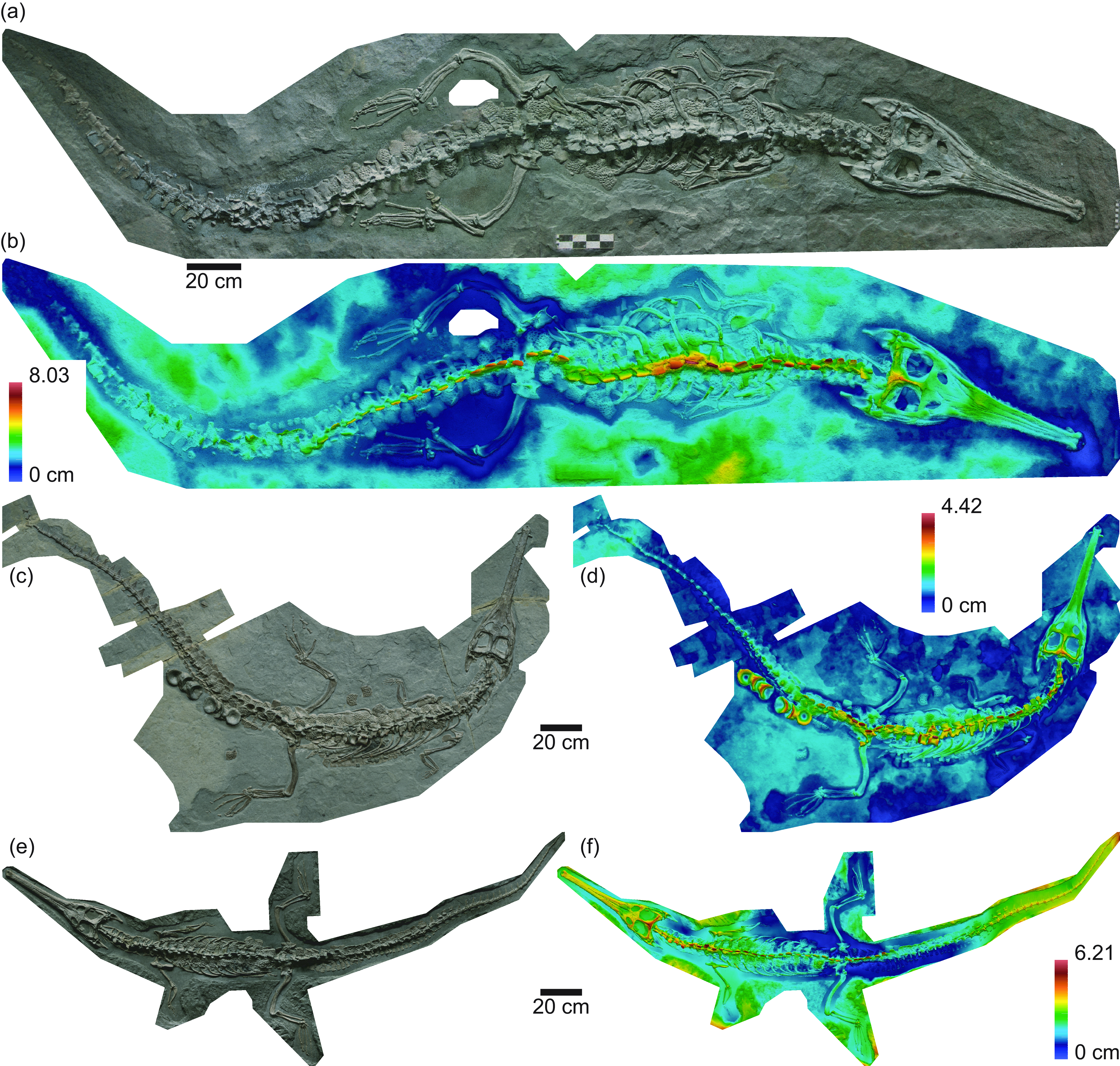
Figure 5. Features of non-headfirst seafloor arrival teleosauroids. Orthophotos of the 3D models of SMNS 54603 (a, b), SMNS 51984 (c, d) and SMNS 52475 (e, f), including images with original texture (a, c, e) and false-colour depth maps (‘Rainbow desaturated’) (b, d, f). Note that SMNS 52475 was originally modelled by H. Mallison and recently modified by E.M.
Only general ventral landings (SMNS 51984, SMNS 52475; or slightly right ventrolateral: SMNS 54603) are interpreted in association with non-headfirst seafloor arrivals in the modelled specimens (Table 2).
4.c. Frequency of headfirst seafloor arrivals
In addition to the thirteen specimens analysed and 3D-modelled (representing ten headfirst and three non-headfirst seafloor arrivals), we examined in-person and/or via photographs 28 additional teleosauroid specimens to determine if headfirst seafloor arrivals are more commonly found than non-headfirst seafloor arrivals. We also tested whether the features described above to identify whether a specimen represents a headfirst or a non-headfirst seafloor arrival could be extrapolated in the absence of 3D models. Our results show that the set of features for each type of arrival can be identified regardless of the orientation of the carcass with respect to the substrate (ventral, dorsal, etc.), as well as the preparation side (from the top- or underside; see Discussion below). Our findings are summarized in Table 2 and specimen photographs are provided in Supplementary Figure 6.
Interestingly, out of 41 total specimens (13 modelled and 28 additional; Table 2), the overall total number of headfirst and non-headfirst seafloor arrivals is approximately equal, being 20 and 21, respectively. Regarding the orientation of the individuals when landing, our interpretations are as follows: five are interpreted as left dorsolateral landings; four as right dorsolateral landings; six as general dorsal landings; four as right ventrolateral landings; six as left ventrolateral landings; fifteen as general ventral landings; and one unknown (Table 2). Only four non-headfirst seafloor arrival specimens (OUMNH JZ176, PIN 2420/1, SMNS 59736 and UMH 0002) are not interpreted as ventral landings (Table 2).
5. Discussion
As noted above, numerous unambiguous features are associated with a headfirst seafloor arrival carcass (Table 1). It is important to reiterate that not all of them are visible or present in all examined specimens (which may be due to different biological, environmental and/or preparation influences), but that the presence of at least one of these features is sufficient to identify the specimen as a headfirst seafloor arrival individual, regardless of inferred biological age stage, size and/or environmental factors. Intertemporal bar inclination and pterygoid perimortem displacement and/or fracturing are commonly seen in conjunction with at least one other feature. In addition, our definition of what constitutes a headfirst seafloor arrival (see Introduction above) exclusively includes specimens in which it can be determined that the actual ‘head’ of the animal landed first and can be applied to not only thalattosuchian crocodylomorphs but also other marine vertebrates as well. It is important to note that specimens identified as non-headfirst seafloor arrivals (e.g. SMNS 54603) may have still impacted the substrate with the ‘head’ (i.e. the skull) first but did not experience any actual damage to the skull and neck. Such a situation can be envisioned if (1) the sediment was very soft, (2) the carcass landed at a low speed (insufficient to create damage at the moment of seafloor impact) and/or (3) an individual died low in the water column. It is a possibility that non-headfirst seafloor arrival specimens may have only landed (nearly) totally ventrally on the substrate, but this is nearly impossible to determine. Moreover, understanding how carcass embedding might be related to taphonomic deformation can inform more accurate morphological interpretation (e.g. whether the size and shape of the orbit or upper temporal fenestra have been altered).
5.a. Interpretation of historical specimens
The interpretation of how each individual specimen was prepared from the surrounding matrix was critical in assessing whether it was representative of a headfirst or non-headfirst seafloor arrival. Large marine vertebrate specimens from the Posidonienschiefer Formation have historically been prepared from either the topside or the underside (stratigraphically upper or lower surface, respectively), and whether a specimen has been prepared from the top or the bottom can have drastically different interpretations and influence how the elevation or depth maps obtained from the 3D photogrammetric models are examined (e.g. Maxwell et al. Reference Maxwell, Cooper, Mujal, Miedema, Serafini and Schweigert2022). Unfortunately, most of the specimens included in our study (Supplementary Table 1) did not have this type of information readily available. As such, we used a critical, logical criterion of observational information gleaned from distinct features (i.e. articulation of elements and their embedding in the sedimentary matrix, see Material and methods above) to determine whether a specimen was prepared either from the topside or underside; in addition, the resulting 3D models were used to strengthen our interpretations regarding the side of preparation. In this regard, we blind-tested our interpretations by comparing them with the information on preparation available from limited specimens. As an example, we initially classified SMNS 51555 as a characteristic headfirst seafloor arrival individual, including features such as a displaced skull and neck, angled fractures in the posterior cranium and mandible and displaced cervical vertebrae (Fig. 3f, Supplementary Fig. 2i, j, Supplementary Table 1). Data from both the skeletal material and 3D photogrammetric model influenced our interpretation that the specimen was prepared from the topside. Fortunately, we then found additional written information on SMNS 51555, which included a note that the specimen had been prepared from the top. As such, the depth information provided by the 3D model was an important and reliable source, as we interpreted SMNS 51555 as prepared from the topside and this agreed with the information associated with the specimen.
Another factor that can influence taphonomic interpretations is dealing with specimens that are imbedded into a different sedimentary layer. For this, the original fossil is mechanically removed and imbedded into a foreign slab of matrix for increased stability and/or aesthetic purposes. This practice is detrimental for reconstructing the original posture and association of the carcass with its original environment, as often during this process disarticulated and/or incomplete skeletons are ‘reconstructed’ or even composited with remains of different individuals. For example, SMNS 15391 (a juvenile Macrospondylus bollensis prepared prior to 1926) was initially included in our headfirst arrival dataset. However, the specimen had been removed from its original surrounding matrix and imbedded in a different layer (Fig. 6), which is clearly seen by examining grain size and colour. The original stratigraphic surface was therefore nearly impossible to observe. Intrinsically, our interpretation of a 3D model for SMNS 15391 would be unreliable, as any stratigraphic plane used would not be the original. In addition, reconstruction or associated plaster casts of skeletal material is another difficulty when considering taphonomic interpretations, as they are not necessarily a reliable source for accurate information. For example, all cervical vertebrae of SMNS 15391 are reconstructed (Fig. 6a); as such, we did not include this specimen for our dataset as many of our headfirst seafloor arrival interpretation(s) reference the neck. Moreover, the false-colour depth map from the 3D model of SMNS 15391 shows that the skull lies at a markedly higher position than the rest of the skeleton (Fig. 6b). We interpret this as the skull being imbedded slightly higher than the rest of the skeleton during preparation. In fact, such difference is not observed in any other false-colour depth map from the other specimens modelled (Figs. 3b, d, 4d, 5b, d, f; Supplementary Figs. 1–4), even from those specimens in which the skull is physically detached from the postcranium (e.g. SMNS 52033) and/or there is a clear displacement of the cervical vertebrae (e.g. SMNS 56370). In combination with the reconstructed cervical region, this suggests extensive modification of SMNS 15391 during the preparation and positioning of the fossil into the new slab.
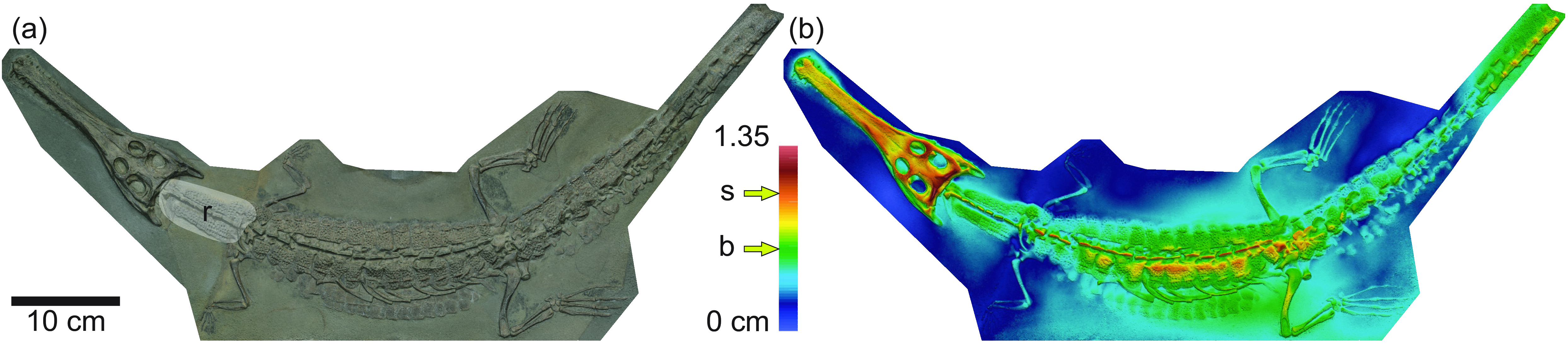
Figure 6. 3D model in orthogonal view of SMNS 15391, a juvenile individual reassembled in foreign sedimentary slabs during preparation. (a) Image with original texture; note that the neck (region shaded) is reconstructed (‘r’). (b) False-colour depth map (‘Rainbow desaturated’); note that the skull is in a higher position than the rest of the body (average positions arrowed in the colour-height scale with ‘s’ and ‘b’, respectively).
5.b. Body size
The total body size and weight of an animal may affect how it moves, the speed at which it moves and how it interacts with the surrounding environment (Webb Reference Webb1984; Massare Reference Massare1988). This is particularly true for marine taxa, as they reside within a unique habitat and thus deal with an exclusive set of environmental, physical and physiological parameters (e.g. Webb Reference Webb1984, Reference Webb1988; Fish Reference Fish2000; Kelley & Motani Reference Kelley and Motani2015; Gutarra & Rahmen Reference Gutarra and Rahman2022). As such, the overall size and body weight, as well as shape, of marine animals is a major influence on how they can impact the seafloor.
Due to a greater size, and subsequently a greater mass, it is possible that subadult (= large skeletally immature individuals) and adult (= large skeletally mature individuals; see Johnson et al. Reference Johnson, Amson and Maxwell2023, Reference Johnson, Scheyer, Canoville and Maxwell2024) teleosauroids were particularly susceptible to skeletal consequences of a headfirst seafloor arrival, whereas in smaller individuals these features are less obvious (e.g. SMNS 10000; Fig. 4c, d). Since smaller specimens are scarcer in the Posidonienschiefer Formation, evaluating taphonomic features to identify potential patterns on preservation is more difficult. Nevertheless, SMNS 10000 is a well-articulated juvenile individual with the skull and mandible stratigraphically lower than the rest of the skeleton, a feature only clearly visible in the false-colour depth map of its 3D model (Fig. 4d). This indicates that the head landed first and penetrated the substrate at a certain angle. Despite the obvious impact that the head and neck underwent, no other features that evidence headfirst seafloor arrival in adult individuals are observed. This supports our hypothesis that such features are less obvious in smaller individuals, and thus, that lower mass of the body did not substantially damage either the skull or neck when the head impacted first on the substrate. Additional specimens are necessary to further strengthen this hypothesis.
5.c. Velocity
It has been hypothesized that teleosauroids used a similar mode of locomotion as modern crocodylians (Massare Reference Massare1988), consisting of axial subundulatory swimming (i.e. small body undulations and sweeping tail undulations, limbs passively held at the sides) and slower speeds (Manter Reference Manter1940; Lighthill Reference Lighthill1969; Frey Reference Frey1982; Fish Reference Fish1984; Braun & Reif Reference Braun and Reif1985). Data provided by Massare (Reference Massare1988) suggested that teleosauroids were not optimally efficient swimmers and were slower than most other Mesozoic marine reptiles (with a calculated speed of approximately 2.2 m/sec.). However, the hypothesis of von Huene (Reference Huene1922), postulating that injured or disoriented animals actively swam headfirst into the seafloor, lacks evidentiary support (e.g. Wetzel & Reisdorf Reference Wetzel, Reisdorf, Bromley, Buatois, Mángano, Genise and Melchor2007), and it is more likely that the animals were dead prior to impact. This means that impact force was determined by body mass and acceleration of the sinking carcass through the water column.
As noted for ichthyosaurs (Reisdorf et al. Reference Reisdorf, Bux, Wyler, Benecke, Klug, Maisch, Fornaro and Wetzel2012), the proximal cause of death for most thalattosuchians not instantly killed by external forces was drowning and is expected to represent the majority of the fossil sample. The large, heavily ossified skull, osteoderm cover and frequent presence of gastroliths, when combined with lungs full of water rather than air almost certainly resulted in a carcass sinking rapidly through the water column. However, the velocity a 3-meter-long teleosauroid needed to achieve within the water column to land headfirst into the sediment with resulting cranial fracturing and vertebral displacement is unclear. Such an analysis is beyond the scope of the current project but will be evaluated in a separate work by the authors in the future.
5.d. Substrate and environment
The Posidonienschiefer Formation is a well-known pelagic (open ocean) environment which has been interpreted as being deposited in an offshore epicontinental setting associated with multiple intervals of benthic anoxia (e.g. Kaufmann Reference Kaufmann1978, Reference Kauffman, Gray, Boucot and Berry1981; Ziegler Reference Ziegler1982; Etter & Tang Reference Etter, Tang, Bottjer, Etter, Hagadorn and Tang2002; Frimmel et al. Reference Frimmel, Oschmann and Schwark2004; Galasso et al. Reference Galasso, Schmid-Röhl, Feist-Burkhardt, Bernasconi and Schneebeli-Hermann2021; Muscente et al. Reference Muscente, Vinnes, Sinha, Schiffbauer, Maxwell, Schweigert and Martindale2023). The Posidonienschiefer Formation in the Southwest German Basin is influenced by a marine transgression whereby the lower parts of this formation (Tenuicostatum Zone) represent a comparatively shallower water environment deposited closer to the paleo-shoreline, but water depth progressively deepens up-sequence with the setting shifting basin-wards towards the top of the section (Bifrons Zone) (Röhl et al. Reference Röhl, Schmid-Röhl, Oschmann, Frimmel and Schwark2001; Röhl & Schmid-Röhl Reference Röhl and Schmid-Röhl2005). Precise water depth values for the Posidonienschiefer Formation in the Holzmaden area remain ambiguous; estimates based on the Dotternhausen locality place the upper Tenuicostatum Zone possibly as shallow as 50 metres, deepening to up to 150 m in the upper Serpentinum Zone (Riegraf et al. Reference Riegraf, Werner and Lörcher1984; Röhl et al. Reference Röhl, Schmid-Röhl, Oschmann, Frimmel and Schwark2001). The lithology is composed of very fine-grained siliciclastic and bituminous claystones, which are well stratified, laminated at some horizons, contain high quantities of pyrite and often lack trace fossils or epibenthos, suggesting a restricted environment of deposition at the seafloor. These quiet conditions, with the redox boundary fluctuating near the sediment-water interface (Muscente et al. Reference Muscente, Vinnes, Sinha, Schiffbauer, Maxwell, Schweigert and Martindale2023), are likely associated with the exceptional preservation of the fossils found in these layers, which include fully articulated vertebrate skeletons with associated soft tissues (Keller Reference Keller1992; Martill Reference Martill1993; Urlichs et al. Reference Urlichs, Wild and Ziegler1994; Lindgren et al. Reference Lindgren, Sjövall, Thiel, Yheng, Ito, Wakamatsu, Hauff, Kear, Engdahl, Alwmark, Eriksson, Jarenmark, Sachs, Ahlberg, Marone, Kuriyama, Gustafsson, Malmberg, Thomen, Rodríguez-Meizoso, Uvdal, Ojika and Schweitzer2018).
The upper layers of the sediment were soft, which allowed vertebrate carcasses to rapidly sink and be partially entombed upon arrival at the seafloor. Carcasses are often asymmetrically articulated, with the side of the carcass originally exposed to the environment on the seafloor being disarticulated as a result (Hofmann Reference Hofmann1958; Martill Reference Martill1993). Substrate consistency likely varied through time in the Holzmaden region. Low calculated sedimentation rates in the lower Serpentinum Zone (Ruebsam et al. Reference Ruebsam, Schmid-Röhl and Al-Husseini2023), in association with a large sample of relatively well-articulated vertebrates, something indicative of limited carcass seafloor exposure time, appear to support a very soft substrate (Martill Reference Martill1993). However, other horizons such as the ‘Schlacken’ (ԑII12) show evidence of benthic infauna, associated with winnowed laminae of eroded bioclasts and/or short-lived event horizons (comprised mostly of small fish bones). Bedding laminae are poorly preserved and most vertebrates are highly disarticulated (Riegraf et al. Reference Riegraf, Werner and Lörcher1984). These horizons therefore likely represent environments with much firmer substrates, resulting in reduced initial burial depth, longer seafloor exposure times and greater degrees of disarticulation for vertebrate carcasses.
The slow sedimentation rates calculated for the Posidonienschiefer Formation (Ruebsam et al. Reference Ruebsam, Schmid-Röhl and Al-Husseini2023), and particularly for the uppermost beds of the Serpentinum Ammonite Zone which produces the majority of the thalattosuchian skeletons (Hauff Reference Hauff1921; Riegraf et al. Reference Riegraf, Werner and Lörcher1984; Röhl & Schmid-Röhl, Reference Röhl and Schmid-Röhl2005; Ruebsam et al. Reference Ruebsam, Schmid-Röhl and Al-Husseini2023), suggest that imbedding depth played a critical role in the high degree of skeletal articulation observed in this konservat lagerstätte. Upon arrival at the seafloor, specimens would have become embedded to a depth proportional to sediment consistency, descent angle, carcass mass, carcass velocity at time of impact and carcass orientation at impact; these factors have been suggested from fish specimens from the Lower Cretaceous Santana Formation of northeastern Brazil (Martill, Reference Martill1997). Given the inferred water depth and bottom temperature at the time of deposition, gas build-up during decomposition would not have refloated the carcasses (Reisdorf et al. Reference Reisdorf, Bux, Wyler, Benecke, Klug, Maisch, Fornaro and Wetzel2012). Articulation and completeness data support the conclusion that decomposition, disarticulation and element loss in the Posidonienschiefer Formation thalattosuchians occurred while the specimens were exposed on the seafloor (Beardmore et al. Reference Beardmore, Orr, Manzocchi and Furrer2012a).
5.e. Predation
Perimortem trauma, scavenging, pathology and associated deadfall communities are often linked with loss of skeletal completeness and disarticulation in marine vertebrates (e.g. Meyer Reference Meyer1991; Harper et al. Reference Harper, Forsythe and Palmer1998; Schmeisser McKean & Gillette Reference Schmeisser McKean and Gillette2015; Delsett et al. Reference Delsett, Novis, Roberts, Koevoets, Hammer, Druckenmiller and Hurum2016; Klompmaker et al. Reference Klompmaker, Kelley, Chattopadhyay, Clements, Huntley and Kowalewski2019; Bosio et al. Reference Bosio, Collareta, Di Celma, Lambert, Marx, de Muizon, Gioncada, Gariboldi, Malinverno, Varas-Malca, Urbina and Bianucci2021; Maxwell et al. Reference Maxwell, Cooper, Mujal, Miedema, Serafini and Schweigert2022; Spiekman & Mujal Reference Spiekman and Mujal2023; Scheyer et al. Reference Scheyer, Johnson, Bastiaans, Miedema, Maxwell and Klug2024), which could theoretically bias how we interpreted imbedding position of the carcasses. However, we found no evidence of trophic interaction related to scavenging (such as bite marks) or deadfall association that can be associated with the landing of the carcasses of any of our 3D-modelled SMNS specimens. Most likely, the specimens in our study identified as headfirst seafloor arrivals correspond to carcasses that were intact prior to landing and remained relatively undisturbed by large scavengers after landing. Disarticulation observed in some specimens is attributed to carcass decay (Beardmore et al. Reference Beardmore, Orr, Manzocchi, Furrer and Johnson2012b), although we cannot rule out scavenging activity not resulting in visible modification of bones, and all fractures and damaged bones of the cranium, mandible and neck are attributed to the headfirst landing itself, not to injuries sustained higher in the water column leading to death. Identifying taphonomic causes of perimortem breakage related to angled cranial embedding can test arguments for potential trophic interactions being responsible for skeletal fracturing. For example, SMNS 53422 was described in the public exhibition as a pabulite, but headfirst seafloor arrival better explains carcass disposition and all observed skeletal damage.
5.f. Skeletal comparison with other marine vertebrates
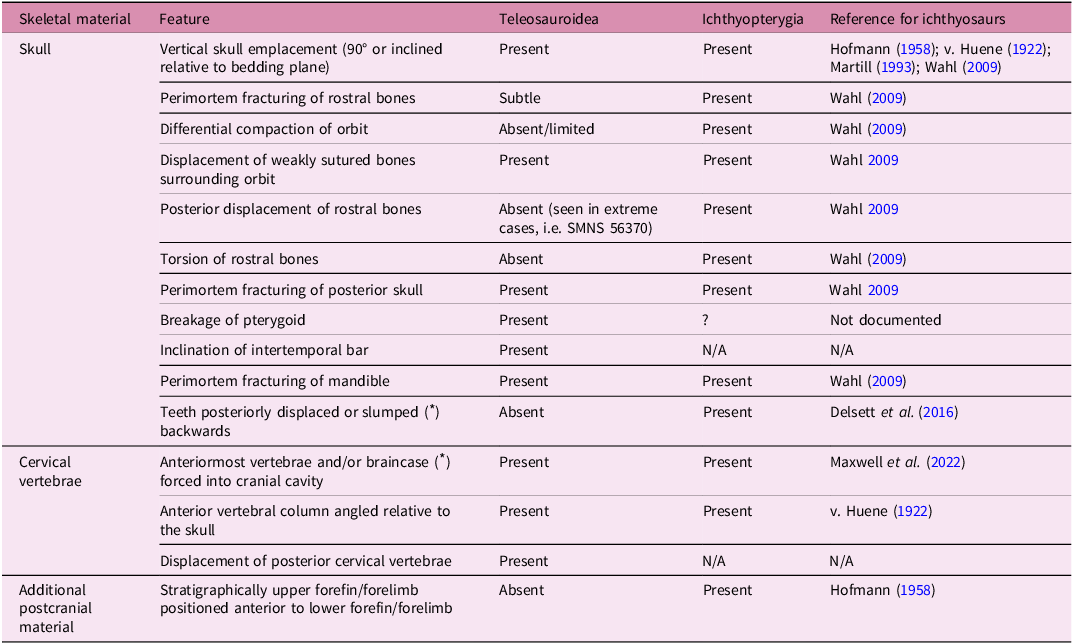
As mentioned above, there are many documented instances of ichthyosaurs exhibiting headfirst seafloor arrivals (Hofmann Reference Hofmann1958; Martill Reference Martill1993; Wahl Reference Wahl2009; Delsett et al. Reference Delsett, Novis, Roberts, Koevoets, Hammer, Druckenmiller and Hurum2016; Maxwell et al. Reference Maxwell, Cooper, Mujal, Miedema, Serafini and Schweigert2022), including specimens from the Posidonienschiefer Formation. As with our teleosauroid sample, distinct features have been associated with headfirst seafloor arrivals in ichthyosaurs. Interestingly, there are multiple cranial and cervical headfirst seafloor arrival features that overlap between teleosauroids and ichthyosaurs (Table 3). These include: (1) skull emplacement that is non-parallel to the substrate, either 90° or inclined to the bedding plane; (2) anterior vertebral column angled relative to the skull; (3) displacement of the skull from the vertebral column; (4) displacement of weakly sutured bones (e.g. jugal) surrounding the orbit; (5) perimortem fracturing of the posterior skull; and (6) forceful intrusion of the braincase and/or anterior cervical vertebrae into the cranial cavity (Table 3). However, aside from the cervical vertebrae (see Table 1), we did not observe any characteristic features in the teleosauroid postcrania that were indicative of a headfirst seafloor arrival; this differs from ichthyosaurs, in which a stratigraphically upper forefin positioned anterior to the lower forefin (Table 3) is associated with a headfirst seafloor arrival (e.g., Fig. 7a, b, Supplementary Fig. 5a, b). In addition, ichthyosaurs impacting the seafloor headfirst display (1) both torsion and displacement of the rostral bones and (2) posterior displacement of the teeth (Table 3), which are both absent in teleosauroids. These points illustrate the role of taxon-specific anatomical differences in the characters used to identify headfirst seafloor arrival. In this regard, the new definition of headfirst seafloor arrival herein provided intends to encompass all the different features observed in different taxonomic groups in order to make it applicable to any marine vertebrate (see also below).
Table 3. Characteristic features (presence of absence) of headfirst seafloor arrivals in Posidonienschiefer Formation teleosauroids and ichthyosaurs
(*) not explicitly stated in the cited reference.

Figure 7. Headfirst seafloor arrivals in other vertebrates. (a, b) Orthophotos of the 3D model of SMNS 56856 (Stenopterygius quadriscissus). (c, d) Orthophotos of the 3D model of SMNS 51199 (Saurostomus esocinus). Both specimens are prepared from the underside. The images are with original texture (a, c) and false-colour depth map (‘Rainbow desaturated’) (b, d). Note that SMNS 56856 was originally modelled by D. Aubert and recently modified by E.M.
It has been proposed that the ‘head-down vertical imbedding orientation’ of some ichthyosaur skulls is restricted to thunniform animals (Wetzel & Reisdorf Reference Wetzel, Reisdorf, Bromley, Buatois, Mángano, Genise and Melchor2007). However, our results show that this is not restricted to animals with a thunniform body shape (e.g. ichthyosaurs) but is also present in elongated bodies with a flexible neck (e.g. teleosauroids).
It is important to note that imbedding and sinking orientation are not synonymous. In parvipelvian ichthyosaurs, in which the neck and torso were relatively inflexible (Lingham-Soliar & Plodowski Reference Lingham-Soliar and Plodowski2007), drag exerted on the carcass would result in a single preferred sinking posture with the skull oriented downwards. In thalattosuchians (excluding more derived metriorhynchids), with a hypothesized laterally flexible neck (Hua Reference Hua2003; Molnar et al. Reference Molnar, Pierce, Bhullar, Turner and Hutchinson2015; Iijima & Kubo Reference Iijima and Kubo2019; Gutarra et al. Reference Gutarra, Stubbs, Moon, Heighton and Benton2023) and a proportionately heavier torso due to the presence of osteoderms and gastroliths, sinking positions might have been less constrained. Thus, headfirst imbedding in thalattosuchians might not resemble the Martill (Reference Martill1993) model for thunniform animals but be more similar to the head-down-body horizontal posture described and illustrated by Martill (Reference Martill1987). Headfirst seafloor arrival has not yet been reported for sauropterygians (Delsett et al. Reference Delsett, Novis, Roberts, Koevoets, Hammer, Druckenmiller and Hurum2016).
Headfirst arrival is not restricted to marine reptiles, as several large-bodied fishes in the Posidonienschiefer Formation, notably the fusiform asthenocormine pachycormiforms (see Cooper et al., Reference Cooper, Giles, Young and Maxwell2022), also exhibit evidence for this imbedding posture. Several large specimens of Saurostomus esocinus preserve the skull at a lower stratigraphic level than the rest of the skeleton, with the posterior portions of the axial skeletons (which were originally exposed on the seafloor) often highly disarticulated or missing (Fig. 7c, d, Supplementary Fig. 5c, d; see also Cooper & Maxwell Reference Cooper and Maxwell2022, Figs. 6b, 6d, 7a). The streamlined nature and torpedo shape of these fishes undoubtedly allowed them to sink at a high velocity, possibly close to a vertical orientation similar to that inferred for thunniform marine reptiles (sensu Wetzel & Reisdorf Reference Wetzel, Reisdorf, Bromley, Buatois, Mángano, Genise and Melchor2007; Cooper & Maxwell Reference Cooper and Maxwell2022). In these specimens, the pectoral fins are laterally splayed and also imbedded in a lower stratigraphic level than the rest of the skeleton (Lingham-Soliar & Plodowski Reference Lingham-Soliar and Plodowski2007). From this, we interpret the extended pectoral fins likely acted as wing-like hydrofoils, reducing lateral tilting and causing the carcass to land headfirst in a ventral-down orientation; this has also been observed in Cretaceous fishes from the Santana Formation (Martill, Reference Martill1997). The force of the impact would have imbedded the skull and pectoral fins deeper in the soft sediment compared to the posterior portion of the fish (Fig. 7d; Cooper et al. Reference Cooper, Giles, Young and Maxwell2022). Other individuals can also display a distinct sigmoidal anterior distortion of the vertebral column (Cooper & Maxwell Reference Cooper and Maxwell2022, Fig. 5b) and in rare cases, a broken rostral bone (rostrodermethmoid) which may have been damaged during a forceful headfirst seafloor arrival (pers. obs. SLAC). Actinopterygian fishes are morphologically dissimilar from ichthyosaurs and thalattosuchians, and therefore, the anatomical criteria outlined here for headfirst seafloor arrival cannot be uncritically applied to fossil fishes. Nonetheless, the new definition for headfirst seafloor arrival here provided still applies to non-reptilian marine vertebrates.
Overall, the new definition of headfirst seafloor arrival of vertebrates that we provide in this work (see Introduction above) intends to integrate all the aforementioned features and points, and it is thus applicable to any marine vertebrate. Yet, the preceding events and consequences of a headfirst landing may differ between taxonomic groups, and even among individuals from the same group. In this regard, our concept of headfirst seafloor arrival is inclusive and describes a set of instances that led to peculiar, yet widespread and relatively diverse, taphonomic pathways of marine vertebrates.
6. Conclusions
Here, we provide the first evidence for headfirst seafloor arrival in thalattosuchian crocodylomorphs, based on a sample of 41 specimens from the Lower Jurassic Posidonienschiefer Formation. We define headfirst landing as: ‘individual(s) that display unambiguous skeletal evidence of the head preceding any other anatomical region in penetrating or landing on the seafloor.’ This definition is applicable not only to teleosauroids but also to other marine vertebrates independent of anatomy.
We provide a comprehensive list of features, mainly pertaining to the skull and neck, that can be attributed to either headfirst or non-headfirst seafloor arrivals in teleosauroids, as well as seafloor landing interpretations. These indicators include features related to either (a) fractures caused by forces applied parallel to the long axis of the skull, as in an anterior collision with the seafloor or (b) unequal taphonomic deformation related to the skull becoming embedded at an angle relative to the bedding plane. While some characteristics are similar to those defined for other marine reptiles (ichthyosaurs) in previous studies, there are taxon-specific differences related to anatomy and body plan. Approximately half of the teleosauroids in our sample (20 out of 41) show osteological and positional indicators characteristic of this landing posture. We also recommend exercising caution when working with historical specimens that have been reset or extensively reconstructed, as how a specimen is prepared can have drastic influences on interpretations.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/S0016756825100058
Data availability statement
Data available from the Figshare Repository: DOI: 10.6084/m9.figshare.29329895.
Acknowledgements
We would like to thank I. Werneberg and A. Krahl (GPIT), M. Wilmsen (MMG), U. Göhlich (NHMW), H. Ketchum (OUMNH) and R. and F. Hauff (UMH) for access to collections, and W. Simpson (FMNH), S. Sachs (NAMU), S. Scherrer (NMWIN), A. Sennikov (PIN), B. Kear (PMU) and G. Wahlefeld and A. Matzke (RNHM) for specimen photographs and information. We sincerely thank H. Mallison for the 3D models of SMNS 9930, SMNS 51563 and SMNS 52475 and D. Aubert for preparing the model of SMNS 56856. We thank L. Ferrari (SMNS) for assistance in the processing of some of the 3D models. We thank David Martill and an anonymous referee, whose reviews helped to improve a previous version of the manuscript, as well as the editor Dr. Bas Van de Schootbrugge for handling the manuscript. This work was supported by the Alexander von Humboldt Foundation (M.M.J., grant number CAN 1218253 HFST-P) and the Deutsche Forschungsgemeinschaft (S.L.A.C. and E.E.M., grant number DFG MA 4693/7-1).
Competing interests
The authors declare none.













 Open access
Open access