


Introduction
Mountain regions are complex ecosystems of conservation interest, harbouring significant biological diversity, and are considered biodiversity hotspots at the global scale (Myers et al. Reference Myers, Mittermeier, Mittermeier, Da Fonseca and Kent2000), with high rates of endemism (Antonelli et al. Reference Antonelli, Kissling, Flantua, Bermúdez, Mulch and Muellner-Riehl2018; Körner et al. Reference Körner, Jetz, Paulsen, Payne, Rudmann-Maurer and Spehn2017). Although mountainous regions cover only 25% of the terrestrial land area, they host 87% of the world’s bird, amphibian, and mammal species, including the majority of species from these taxa with restricted geographical ranges (Rahbek et al. Reference Rahbek, Borregaard, Colwell, Dalsgaard, Holt and Morueta-Holme2019). Mountains have a deep cultural importance for human populations, who have significantly shaped these ecosystems and their associated biodiversity through various activities such as agriculture and forestry, and more recently, for renewable energy and leisure activities.
The distribution and population trends of birds in Europe have been influenced over time by anthropogenic activity (BirdLife International 2021; Keller et al. Reference Keller, Herrando, Voříšek, Franch, Kipson and Milanesi2020). In mountain regions, human activities impacting birds, whether directly or indirectly, are numerous. Among the primary drivers of population decline for mountain birds are hunting, persecution, energy production, and human disturbance (Alba et al. Reference Alba, Kasoar, Chamberlain, Buchanan, Thompson and Pearce-Higgins2022), with snow sports such as skiing being a key source of disturbance. The rapid growth of tourism in the Alps has negatively impacted wildlife (Ingold Reference Ingold2005), especially birds (Caprio et al. Reference Caprio, Rolando, Arlettaz, Chamberlain, Chamberlain, Lehikoinen and Martin2023), primarily due to habitat loss, fragmentation, and human disturbance (Arlettaz et al. Reference Arlettaz, Patthey, Baltic, Leu, Schaub and Palme2007; Larson et al. Reference Larson, Reed, Merenlender and Crooks2019). Furthermore, winter tourism in Europe is expanding, particularly into higher elevations due to climate change (Caprio et al. Reference Caprio, Rolando, Arlettaz, Chamberlain, Chamberlain, Lehikoinen and Martin2023; Marty Reference Marty, Rixen and Rolando2013). Ski tourism harms various animal taxa, including birds, through direct processes such as increased stress (Arlettaz et al. Reference Arlettaz, Patthey, Baltic, Leu, Schaub and Palme2007), decreased survival due to collision with infrastructures (Buffet and Dayot Reference Buffet, Dumont-Dayot, Rixen and Rolando2013), population decrease (Patthey et al. Reference Patthey, Wirthner, Signorell and Arlettaz2008), habitat degradation (Braunisch et al. Reference Braunisch, Patthey and Arlettaz2011), and through broader processes affecting species richness and diversity (Caprio et al. Reference Caprio, Chamberlain, Isaia and Rolando2011; Laiolo and Rolando Reference Laiolo and Rolando2005). The construction and management of ski-pistes cause significant soil damage, with machine grading, artificial snow, and chemicals disrupting soil structure and microbial communities (Burt and Rice Reference Burt and Rice2009; Gros et al. Reference Gros, Monrozier, Bartoli, Chotte and Faivre2004; Roux-Fouillet et al. Reference Roux-Fouillet, Wipf and Rixen2011). However, more sustainable management practices can reduce these impacts, with restored ski-pistes showing higher species richness and abundance compared with unrestored areas (Caprio et al. Reference Caprio, Chamberlain and Rolando2016).
Ski-pistes, which maintain significant amounts of snow through the spring due to the provisioning of artificial snow and snow compaction, might play a role in the feeding ecology of mountain birds (Antor Reference Antor1995; Resano-Mayor et al. Reference Resano-Mayor, Korner-Nievergelt, Vignali, Horrenberger, Barras and Braunisch2019). Arthropod fallout is a snow-related phenomenon described by many authors (e.g. Edwards and Banko Reference Edwards and Banko1976; Papp Reference Papp1978; Spalding Reference Spalding1979), which occurs when flying insects land on the snow surface and become moribund, thus becoming prey for birds. In the 1960s, researchers began observing mountain birds foraging on snow and hypothesised that snow patches could serve as a food source due to this fallout (Pattie and Verbeek Reference Pattie and Verbeek1966). Verbeek (Reference Verbeek1970) experimentally tested this “snowfield hypothesis” by studying feeding rates of birds on snow versus vegetation. His findings showed higher feeding rates on snow. However, subsequent studies revealed mixed results (Antor Reference Antor1995; Norvell and Creighton Reference Norvell and Creighton1990; Zamora Reference Zamora1990). While questions arose about the consistency of arthropod fallout and bird preferences for snow-covered versus snow-free habitats, one consistent finding was that prey detectability and accessibility were greater on snow (Antor Reference Antor1995; Hendricks Reference Hendricks1987; Zamora Reference Zamora1990). Overall, the evidence thus suggests that snowfields are important for mountain birds, especially following high winds that cause substantial arthropod fallout (Brambilla et al. Reference Brambilla, Capelli, Anderle, Forti, Bazzanella and Masiero2018; Papp Reference Papp1978).
As climate change threatens open-habitat mountain species and their habitats (Brunetti et al. Reference Brunetti, Lentini, Maugeri, Nanni, Auer and Bohm2009; IPCC 2013), studies on the foraging preferences of mountain birds have increased, highlighting the continued relevance of snow and snow-associated habitats (e.g. Alessandrini et al. Reference Alessandrini, Scridel, Boitani, Pedrini and Brambilla2022; Brambilla et al. Reference Brambilla, Cortesi, Capelli, Chamberlain, Pedrini and Rubolini2017; Müller et al. Reference Müller, Meier, Knaus, Korner, Helm and Amrhein2023; Resano-Mayor et al. Reference Resano-Mayor, Korner-Nievergelt, Vignali, Horrenberger, Barras and Braunisch2019). These habitats are not only important for arthropod fallout, but also for the emergence of insect larvae in spring, particularly along melting snow patch margins where insects become concentrated as snow recedes. Despite this, no study has yet examined the role of these dynamic microhabitats in the feeding ecology of mountain birds at the community level, with existing research primarily focused on single species such as White-winged Snowfinch Montifringilla nivalis (e.g. Brambilla et al. Reference Brambilla, Cortesi, Capelli, Chamberlain, Pedrini and Rubolini2017, Reference Brambilla, Scridel, Sangalli, Capelli, Pedrini and Bogliani2019; Resano-Mayor et al. Reference Resano-Mayor, Korner-Nievergelt, Vignali, Horrenberger, Barras and Braunisch2019; Scridel et al. Reference Scridel, Anderle, Capelli, Forti, Bettega and Alessandrini2024), Northern Wheatear Oenanthe oenanthe (e.g. Muller et al. Reference Müller, Meier, Knaus, Korner, Helm and Amrhein2023), Ring Ouzel Turdus torquatus (e.g. Barras et al. Reference Barras, Marti, Ettlin, Vignali, Resano-Mayor and Braunisch2020) or choughs Pyrrhocorax spp. (e.g. Rolando et al. Reference Rolando, Laiolo and Formica1997).
Here, we focused on ski-pistes, where the persistence of snow margins can enhance food availability for birds. Due to the compaction of snow and artificial snowmaking, ski-pistes retain snow longer than other habitats (Rixen and Rolando Reference Rixen and Rolando2013), potentially amplifying their value in periods where there is little snow, but when the vegetation at high elevations has yet to emerge (i.e. early spring). Effective snow management could thus benefit bird communities, mitigating some negative impacts of ski-pistes. Ski-pistes are ideal “laboratories” due to their accessibility, the ability to monitor snow phenology via webcams, and the availability of artificially maintained snow. The aims of this study were to: (1) investigate the role of snow and snowmelt in ski-pistes for bird foraging habitat selection; (2) identify key habitat features influencing bird foraging; (3) propose conservation actions to reduce the impacts of the ski industry on mountain biodiversity, thus maximising the potential advantages and minimising the disadvantages of snow patches on ski-pistes. Although our study primarily explores community-level foraging patterns, we decided to deepen the analysis for a species of particular conservation concern, the Water Pipit Anthus spinoletta, which is a high-priority species for conservation in the Alps due to projected reductions in breeding range in the coming decades (Brambilla et al. Reference Brambilla, Rubolini, Appukuttan, Calvi, Karger and Kmecl2022; Chamberlain et al. Reference Chamberlain, Negro, Caprio and Rolando2013). The species shows a decreasing trend across Europe (BirdLife International 2021; Lehikoinen et al. Reference Lehikoinen, Brotons, Calladine, Campedelli, Escandell and Flousek2019), with recent declines documented in some countries (Knaus et al. Reference Knaus, Antoniazza, Wechsler, Guélat, Kéry and Strebel2018; Rete Rurale Nazionale and Lipu 2024). Given its conservation importance and status as a true mountain specialist, we decided to explore the Water Pipit’s foraging habitat selection in relation to snow on ski-pistes, with the goal of informing more targeted conservation measures.
Methods
Study area and survey design
The study area was located in the western Italian Alps in the High Susa Valley at 44°57′N 6°53′E (Figure 1a) covering elevations between 1,870 m and 2,420 m a.s.l. In the area, there are c.400 km of ski-pistes, distributed over 247 single pistes that are parts of one of the largest ski areas in Europe, the “Via Lattea” (https://www.vialattea.it/en/). The dominant forest tree is the European larch Larix decidua, with Norway spruce Picea abies, Swiss pines Pinus cembra, and other broadleaved trees also present. At higher elevations in the treeline ecotone, shrubs such as common juniper Juniperus communis and alpenrose Rhododendron ferrugineum occur, and alpine grasslands and scree are present above 2,300–2,400 m a.s.l.

Figure 1. The location of the study area in the western Italian Alps (a), where 34 line transects (yellow) were placed on the ski-pistes close to the town of Sestriere (b). Ski-pistes were visited during the spring snow melting period in 2023 (c). When a foraging bird was seen, the 5 m radius around its location was defined as a foraging plot which was paired with a randomly selected pseudo-absence plot in the same general area (d). Pseudo-absence plots were generated randomly at 50 m distance and at a random angle (relative to the observer walking direction along the ski-piste) from each foraging location. (Photographs: ©A. Fragomeni)
Using geographic information systems (GIS) software, QGIS 3.14.15 Pi (QGIS Development Team 2023) and satellite images from Google Earth, we randomly placed 34 transects along the elevational gradient and exclusively on ski-pistes (Figure 1b). The purpose of the transect surveys was to detect foraging birds whose locations were used as the basis for the analysis (see below). Each transect was 200 m long and surveys were carried out in 2023 during the spring period over two visits in the early breeding season: the first was carried out from 12 April to 27 April and the second from 16 May to 24 May, with different snowmelt conditions (Figure 1c). Surveys were carried out both during the morning and afternoon when bird foraging activity is highest (9h00–11h00 and 15h00–17h00; see Alba et al. Reference Alba, Sander, Rosselli and Chamberlain2024).
Bird surveys
To study foraging habitat selection by mountain birds, we used pairs of plots with bird presence and control plots with pseudo-absence (Figure 1d). This method allowed for comparison between a site selected for feeding by birds along with its associated habitat features (hereafter “foraging plot”) of 5 m radius and a site of the same size within the same area that apparently was not chosen (hereafter “pseudo-absence plot”) (see Barras et al. Reference Barras, Marti, Ettlin, Vignali, Resano-Mayor and Braunisch2020; Müller et al. Reference Müller, Meier, Knaus, Korner, Helm and Amrhein2023; Resano-Mayor et al. Reference Resano-Mayor, Korner-Nievergelt, Vignali, Horrenberger, Barras and Braunisch2019). Transects were walked at a slow pace and the observer stopped whenever one or more birds were observed foraging, creating a foraging plot. The abundance and species of birds were noted at this plot. A bird was considered to be feeding if it was seen pecking at the ground, with each peck assumed to correspond to an attempt to capture prey. After five minutes, we immediately collected the microhabitat variables on the plot (see Habitat surveys below). A pseudo-absence plot was then created at a 50 m distance from the foraging plot at a random angle (0–360°) relative to the direction in which the observer was walking along the transect, which was set as 0° in each case. We decided to use 50 m as a standard distance, as it was shown to express foraging habitat selection at the scale of the home-range for mountain birds (Barras et al. Reference Barras, Marti, Ettlin, Vignali, Resano-Mayor and Braunisch2020; Burfield Reference Burfield2002). We checked for possible presence of birds in the pseudo-absence plots and then the habitat features were recorded. Distances were measured with a laser range finder.
Habitat surveys
A set of fine-scale habitat variables were collected within a 5 m radius for both foraging and pseudo-absence plots, capturing the environmental conditions that birds encounter while foraging. We sampled the percentage cover of snow, grass, mud, and rock at ground level, with the total sum adding up to 100%. These variables reflect the small-scale habitat heterogeneity that may influence prey availability and accessibility. Grass height was measured at a randomly selected point within the plot to assess vegetation structure. Additionally, we recorded the distance from the nearest forest edge and elevation. Distances were measured using a laser range finder to ensure accuracy at a fine spatial resolution.
Statistical analysis
To investigate the key habitat elements influencing the foraging habitat selection of mountain birds, we modelled three response variables using generalised linear mixed models (GLMMs). First, we assessed bird occurrence probability (i.e. the probability of any bird using a plot) with a GLMM using a binary response variable (presence/absence) and a logit-link function. Next, we explored the factors affecting total bird abundance and species richness through two additional GLMMs, both specifying a Poisson distribution with a log-link function. In all models, the habitat cover estimates, grass height, and distance to forest were included as fixed effects. We checked for collinearity among predictors by calculating Spearman’s correlation coefficient. Grass cover showed a high correlation (r s >0.7) with snow cover. As the main aim of this study was to investigate the relevance of snow in the foraging habitat selection of birds, we decided to retain snow and discard grass cover from the analyses. To understand which key habitat elements were most important for foraging habitat selection, we carried out model averaging for bird presence, abundance, and species richness. We used “dredge” and “model.avg” functions from the MuMIn package (Barton Reference Barton2023) on the best set of models, defined as those within ∆AIC <2. After community analyses, we fitted a species-specific GLMM using a binary response variable (presence/absence) to model the occurrence probability of the Water Pipit in relation to the same habitat predictors. Prior to analyses, all habitat variables were scaled and centred. We checked for possible non-linear relationships when visual inspection of scatterplots suggested their possible existence. To account for spatial autocorrelation, we included point identity (unique number for each presence/pseudo-absence pair) as a random effect. GLMMs were carried out with the glmmTMB R package (Brooks et al. Reference Brooks, Kristensen, van Benthem, Magnusson, Berg and Nielsen2017). For binomial GLMMs, we assessed goodness-of-fit using area under the curve (AUC), with the “auc” and “roc” functions from the pROC package (Robin et al. Reference Robin, Turck, Hainard, Tiberti, Lisacek and Sanchez2011). For the Poisson models, we used the DHARMa package (Hartig Reference Hartig2022) with the “simulatedResiduals” function. No indications of poor model fit were found. All analyses were performed using R software 4.3.1 (R Core Team 2024).
Results
A total of 167 individual birds belonging to 17 species were recorded foraging on ski-pistes (Figure 2). The total sample size was 73 plots (73 foraging plots and the same number of paired pseudo-absence plots). The most abundant species on ski-pistes was Carrion Crow Corvus corone, followed by Water Pipit. Other common species (≥10 individuals) included Fieldfare Turdus pilaris, White Wagtail Motacilla alba, Common Linnet Linnaria cannabina, and Northern Wheatear (Figure 2).
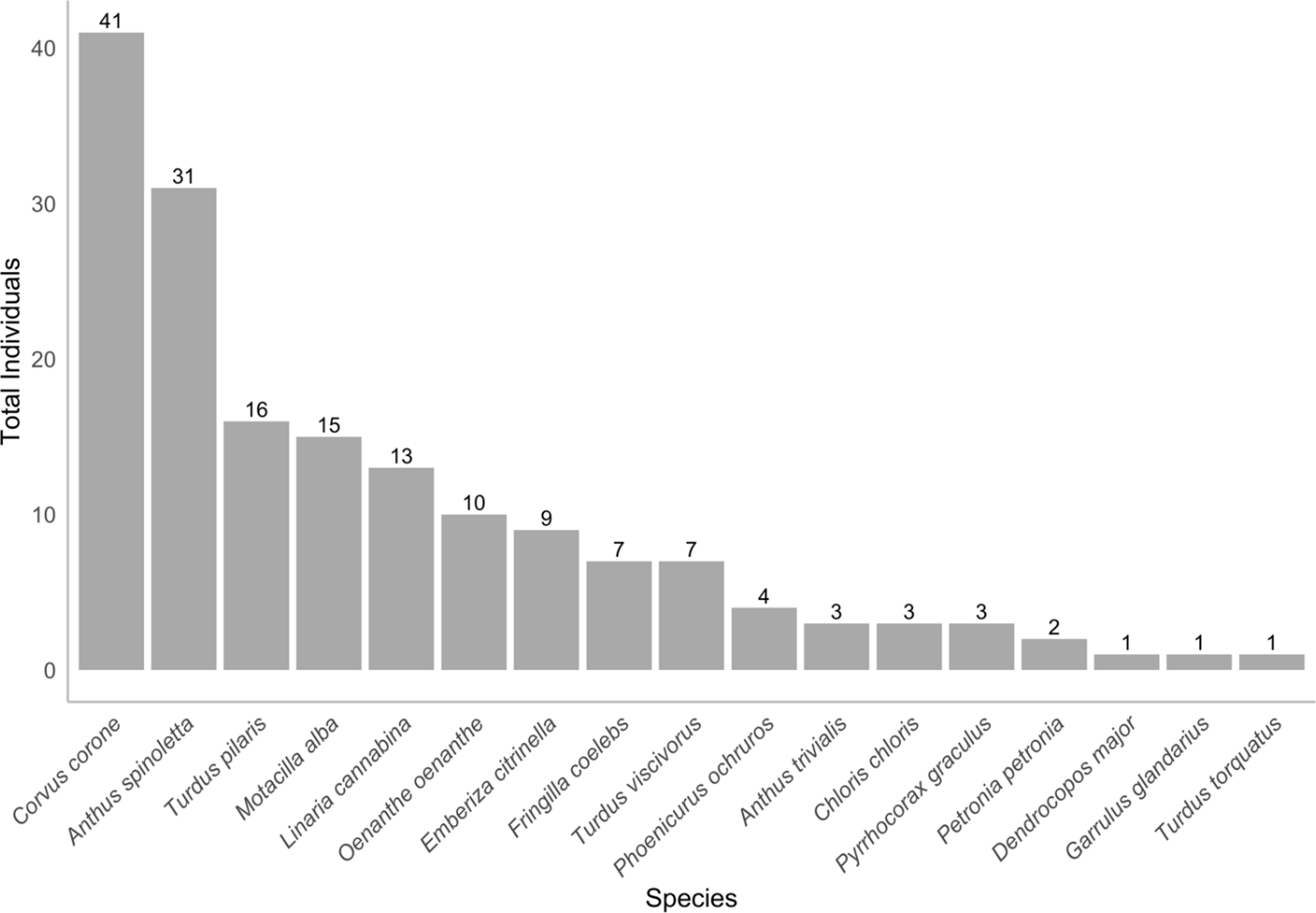
Figure 2. Species and number of individuals contacted during the surveys.
Snow cover showed a significant quadratic effect on occurrence probability, abundance, and species richness, and was the most important variable in all three model sets (Table 1), highlighting the critical role of snow in affecting foraging habitat selection. Mud cover showed a significant positive relationship with occurrence probability, whilst bird abundance significantly decreased with elevation (Figure 3). Full details of model-averaged parameters and variable importance for occurrence, abundance, and species richness models are given in Supplementary material Tables S1 and S2. The occurrence probability of Water Pipit was significantly related to mud cover and, to a lesser extent, grass height, suggesting that areas with greater mud cover and shorter grass height offer more suitable foraging conditions. Furthermore, nearly significant negative relationships with snow cover and distance from forest were highlighted (see Figure 4 and Table S3).
Table 1. Outputs of the final model-averaged parameters for occurrence probability, abundance, and species richness. Estimate, standard error (SE), test value (z), and P value are given for each parameter. Only significant parameters are reported; see variable importance in Table S1 and all parameters in Table S2
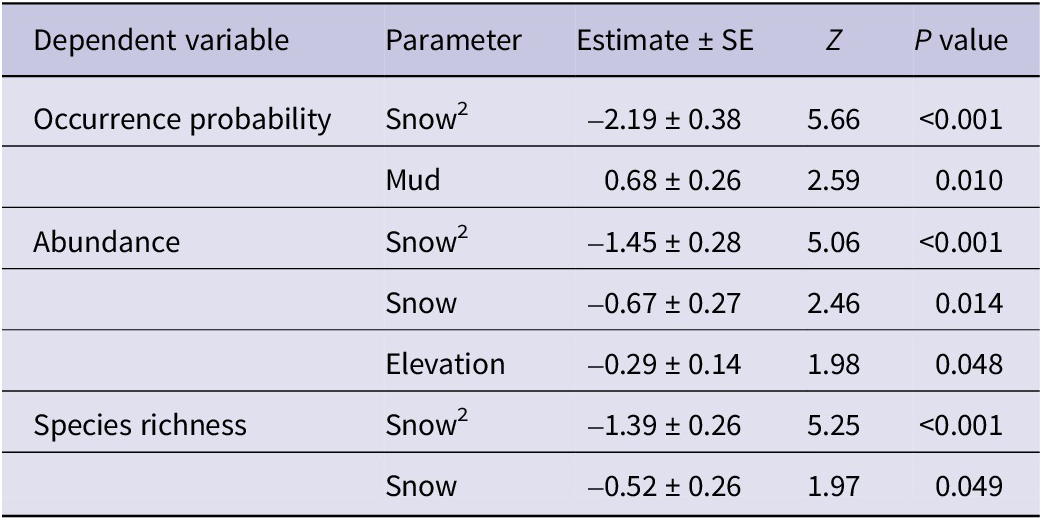


Figure 4. Occurrence probability of the Water Pipit. Black circles represent raw data, whilst P values are shown in the top-right corner of each plot. See all parameters in Table S3.
Discussion
This study underscores the critical role of snow and snow-associated habitats on ski-pistes in the foraging ecology of mountain bird communities. Snow cover emerged as the most influential variable, demonstrating a consistent quadratic effect on bird presence, abundance, and species richness. Birds showed a preference for intermediate snow cover levels, likely due to the habitat heterogeneity and resource availability that these conditions provide. While grass height was not a significant factor in our models, we recognise that its influence might increase later in the season when vegetation height varies more. The findings extend previous species-specific studies, such as those on White-winged Snowfinch (e.g. Alessandrini et al. Reference Alessandrini, Scridel, Boitani, Pedrini and Brambilla2022; Brambilla et al. Reference Brambilla, Cortesi, Capelli, Chamberlain, Pedrini and Rubolini2017; Resano-Mayor et al. Reference Resano-Mayor, Korner-Nievergelt, Vignali, Horrenberger, Barras and Braunisch2019; Strinella et al. Reference Strinella, Ricci and Vianale2007) and Northern Wheatear (Müller et al. Reference Müller, Meier, Knaus, Korner, Helm and Amrhein2023), to a community level, revealing consistent patterns.
The preference for intermediate snow cover likely arises from the enhanced availability and detectability of prey along snow edges. Previous studies have linked arthropod fallout and biomass accumulation to snow margins, particularly for taxa like Tipulidae, Coleoptera, and Araneae, which thrive in moist conditions (Resano-Mayor et al. Reference Resano-Mayor, Korner-Nievergelt, Vignali, Horrenberger, Barras and Braunisch2019). Additionally, it has been shown that some species, such as the Ring Ouzel, prefer habitats with increased levels of moisture while foraging, due to higher soil penetrability (Barras et al. Reference Barras, Marti, Ettlin, Vignali, Resano-Mayor and Braunisch2020). These results align with the observed positive effects of mud cover, often associated with snowmelt. However, while earlier studies have emphasised the role of arthropods as a key resource in snow habitats, they often focused on a narrow subset of species or specific ecological conditions. For instance, Verbeek (Reference Verbeek1970) and Brambilla et al. (Reference Brambilla, Capelli, Anderle, Forti, Bazzanella and Masiero2018) provided foundational insights into snow-associated foraging of individual species, whereas we have provided a comprehensive approach to generalise the findings across the bird community. By encompassing multiple species (and thus including species which were previously not considered in terms of snowmelt habitat exploitation), this study fills an important gap in understanding how snow and its associated features are critical for mountain bird ecology.
Nevertheless, questions remain regarding the consistency of snow-related phenomena such as arthropod fallout. Some earlier studies (e.g. Antor Reference Antor1995; Norvell and Creighton Reference Norvell and Creighton1990) reported variability in prey availability and bird utilisation of snow habitats, often influenced by localised environmental factors like wind conditions or snow quality. Our results suggest that habitat features like mud and snowmelt edges can help create more predictable foraging conditions, although for birds that feed on soil-dwelling invertebrates, rather than those that rely on wind-blown prey. These findings highlight the need to revisit older studies and incorporate updated methodologies, such as remote sensing, to refine our understanding of snow’s ecological role.
The results have significant conservation implications, particularly given the on-going threats to mountain ecosystems from climate, land-use changes, and human disturbance. Ski-pistes, while often viewed as environmentally detrimental (Rixen and Rolando Reference Rixen and Rolando2013), could be managed to provide supplementary foraging habitats for birds. The retention of snow on pistes through artificial snowmaking might help offset the effects of earlier snowmelt due to global warming, extending the availability of these critical habitats into late spring. Similar patterns have been observed in species like the White-winged Snowfinch, which preferentially forages on snow patches and short-sward alpine grasslands found on ski-pistes during the nestling period (Bettega et al. Reference Bettega, Luciani, Roseo, Pedrini, Leonardi and Brambillain press). This species also adjusts foraging behaviour based on temperature and solar radiation, highlighting the complex interplay between microclimate and habitat features.
These findings suggest that piste management strategies, such as reducing machine grading and preserving small snow patches, could benefit not only snowfinches, but a broader spectrum of alpine birds reliant on snow-associated prey. Artificial snow could provide suitable habitat for some bird species in late spring, especially in areas where natural snow melts early due to warmer temperatures. However, it remains a complex and controversial solution. Artificial snowmaking is energy-intensive and contributes to water resource depletion, raising concerns about its long-term sustainability in the context of climate change. Moreover, its impacts on soil structure, vegetation, and microbial communities are still not fully understood (Hudek et al. Reference Hudek, Barni, Stanchi, D’Amico, Pintaldi and Freppaz2020; Rixen et al. Reference Rixen, Stoeckli and Ammann2003), and any potential benefits for birds must be weighed against broader ecological costs. Rather than expanding artificial snowmaking into pristine habitats, a more sustainable approach could involve maintaining snow cover within existing ski areas, particularly in the lower parts of pistes that are already heavily modified. To achieve this, alternative methods to prolong snow presence could be explored, such as creating snow patches or compacted snow “hills” after piste closure, which would melt more slowly due to their volume, or employing snow fences to enhance natural snow accumulation. The usefulness of such techniques in an alpine biodiversity context still needs further exploration. Combining the above strategies with habitat restoration efforts may offer a more environmentally sound trade-off, supporting biodiversity while minimising ecological impacts.
The preference for snow–mud mosaics observed in this study highlights the importance of maintaining habitat heterogeneity on ski-pistes. We showed how the occurrence probability of Water Pipit, a species of conservation concern, was significantly related to mud cover and, to a lesser extent, to grass height, suggesting that areas with greater mud cover and shorter grass height offer more suitable foraging conditions. Conservation strategies should focus on minimising habitat homogenisation caused by grading and other piste-management practices. Restored ski-pistes have been shown to support higher species richness and abundance than unrestored areas (Caprio et al. Reference Caprio, Chamberlain and Rolando2016), suggesting that targeted habitat restoration could enhance the ecological value of ski slopes. Techniques such as re-seeding with native plants, reducing soil compaction, and limiting the use of chemical treatments could help create more bird-friendly environments. The findings also underscore the broader need to address climate change and land-use shifts in mountain ecosystems. Rising temperatures threaten to reduce snow cover and alter the phenology of snowmelt, with cascading effects on alpine biodiversity (Brambilla et al. Reference Brambilla, Cortesi, Capelli, Chamberlain, Pedrini and Rubolini2017). Conservation strategies should include protecting natural snow-associated habitats, such as alpine meadows and pastures, which are crucial for both birds and their invertebrate prey. Policies promoting sustainable agriculture and forestry practices in mountain regions could help maintain these open habitats, complementing efforts to improve ski-piste management.
For instance, ski resorts could be incentivised to adopt biodiversity-friendly practices, such as maintaining buffer zones around pistes and reducing disturbance (e.g. caused by management outside the skiing season) during critical breeding and foraging periods for birds (Lopez et al. Reference Lopez, Chamberlain, Requena, Rosselli and Alba2024). Future research should focus on validating these findings across diverse alpine regions and exploring how snow type (natural vs artificial) influences foraging site selection. Additionally, studies examining the long-term impacts of ski-piste use on bird populations and their prey would provide valuable insights into the sustainability of current management practices.
Conclusions
This study highlights the dual role of snow as both a potential beneficial factor influencing mountain bird feeding ecology and a resource increasingly threatened by human activities and climate change. While ski-pistes can provide foraging habitats under certain conditions, their broader ecological costs cannot be ignored. By adopting more sustainable management practices, ski resorts could help mitigate their environmental impact and contribute to the conservation of mountain biodiversity. However, these efforts must be part of a larger strategy addressing the root causes of land-use changes and climate change in mountain ecosystems. Only through such integrative approaches can we ensure the long-term survival of mountain bird communities and the ecosystems they inhabit.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0959270925100130.








 Open access
Open access