



1. Introduction
Peritrichous bacteria, such as Escherichia coli (E. coli), swim by combining multiple independently driven flagella into a single rotating helical bundle, which generates hydrodynamic thrust and propels the cells forward (Berg Reference Berg2004). While the propulsion mechanism enabled by a single helical bundle is well studied (Lauga Reference Lauga2020), fluid-mediated interactions among multiple flagella within the bundle are still far from fully understood (Bianchi et al. Reference Bianchi, Saglimbeni, Frangipane, Cannarsa and Di Leonardo2023). Although theoretical and numerical modelling have been empolyed to elucidate the collective dynamics of multiple flagella during bundle formation and within a functional bundle (Kim & Powers Reference Kim and Powers2004; Flores et al. Reference Flores, Lobaton, Méndez-Diez, Tlupova and Cortez2005; Reichert & Stark Reference Reichert and Stark2005; Janssen & Graham Reference Janssen and Graham2011; Reigh, Winkler & Gompper Reference Reigh, Winkler and Gompper2012; Kanehl & Ishikawa Reference Kanehl and Ishikawa2014; Man et al. Reference Man, Page, Poole and Lauga2017; Chamolly & Lauga Reference Chamolly and Lauga2020; Tătulea-Codrean & Lauga Reference Tătulea-Codrean and Lauga2021, Reference Tătulea-Codrean and Lauga2022; Park et al. Reference Park, Kim, Lee, Pfeifer, Muraveva, Beta and Lim2024), direct experimental validation of predicted flagellar dynamics remains difficult due to the small spatial scale (
 $\sim 20\,\rm nm$
) and rapid time scale (
$\sim 20\,\rm nm$
) and rapid time scale (
 ${\lt } 10\,\rm ms$
) associated with the flagellar process (Turner, Ryu & Berg Reference Turner, Ryu and Berg2000). To tackle this challenge, scaled experimental models have been developed to mimic flagellar dynamics at macroscopic scales (Macnab Reference Macnab1977; Kim et al. Reference Kim, Bird, Van Parys, Breuer and Powers2003, Reference Kim, Kim, Bird, Park, Powers and Breuer2004; Danis et al. Reference Danis, Rasooli, Chen, Dur, Sitti and Pekkan2019; Lim, Yadunandan & Khalid Jawed Reference Lim, Yadunandan and Khalid Jawed2023). Most of these works, however, focused on the formation of the flagellar bundle. Few experiments have been conducted exploring the hydrodynamic interactions between multiple flagella and examining their effects on the dynamics of flagella in a stable bundle.
${\lt } 10\,\rm ms$
) associated with the flagellar process (Turner, Ryu & Berg Reference Turner, Ryu and Berg2000). To tackle this challenge, scaled experimental models have been developed to mimic flagellar dynamics at macroscopic scales (Macnab Reference Macnab1977; Kim et al. Reference Kim, Bird, Van Parys, Breuer and Powers2003, Reference Kim, Kim, Bird, Park, Powers and Breuer2004; Danis et al. Reference Danis, Rasooli, Chen, Dur, Sitti and Pekkan2019; Lim, Yadunandan & Khalid Jawed Reference Lim, Yadunandan and Khalid Jawed2023). Most of these works, however, focused on the formation of the flagellar bundle. Few experiments have been conducted exploring the hydrodynamic interactions between multiple flagella and examining their effects on the dynamics of flagella in a stable bundle.
Beyond their function in bacterial motility, rotating helices can be harnessed as pumps and mixers in microfluidics. Immobilised bacteria in bacterial carpets generate strong near-surface flows, enabling fluid transport and mixing (Darnton et al. Reference Darnton, Turner, Breuer and Berg2004; Kim & Breuer Reference Kim and Breuer2008), and guiding particle motion along designed trajectories (Gao et al. Reference Gao, Li, Chen and Zhang2015). The possibility of using rotating helices as microfluidic pumps has also been explored in simulations (Martindale & Fu Reference Martindale and Fu2017; Buchmann et al. Reference Buchmann, Fauci, Leiderman, Strawbridge and Zhao2018; Dauparas, Das & Lauga Reference Dauparas, Das and Lauga2018; Rostami et al. Reference Rostami, Liu, Buchmann, Strawbridge and Zhao2022). Artificial helical microrobots actuated by external fields have also been exploited for fluid pumping and mixing (Zhang, Peyer & Nelson Reference Zhang, Peyer and Nelson2010; Tottori et al. Reference Tottori, Zhang, Qiu, Krawczyk, Franco-Obregõn and Nelson2012). These studies demonstrate the great potential of using collectively rotating helices for precise control of low-Reynolds-number flow in microfluidics. A deeper understanding of the hydrodynamic interactions between rotating helices and a detailed mapping of the surrounding flow characteristics become essential to fully realising this potential.
Our work, integrating experiments and hydrodynamic modelling, addresses this critical knowledge gap and technical challenge, and provides an in-depth study of the dynamics of a pair of rigid rotating helices in low-Reynolds-number flows.
2. Methods
2.1. Experimental set-up
We construct a macroscopic model of bacterial flagella. The scaled helical flagella are left-handed and fabricated by wrapping stainless steel wires (radius
 $a = 1\,\rm mm$
) around a cylindrical rod with radius
$a = 1\,\rm mm$
) around a cylindrical rod with radius
 $R = 1\,\rm cm$
at pitch length
$R = 1\,\rm cm$
at pitch length
 $\lambda = 12\,\rm cm$
(figure 1
a). The helical axial length submerged in fluid is fixed at
$\lambda = 12\,\rm cm$
(figure 1
a). The helical axial length submerged in fluid is fixed at
 $L= 32 \,\rm cm$
. The ratios
$L= 32 \,\rm cm$
. The ratios
 $R/L = 0.026$
and
$R/L = 0.026$
and
 $\lambda /L = 0.32$
closely match those of E. coli flagella at
$\lambda /L = 0.32$
closely match those of E. coli flagella at
 $R/L = 0.028$
and
$R/L = 0.028$
and
 $\lambda /L = 0.35$
(Turner et al. Reference Turner, Ryu and Berg2000).
$\lambda /L = 0.35$
(Turner et al. Reference Turner, Ryu and Berg2000).
The schematic of our experimental set-up is shown in figure 1(b–d). The helices are inserted vertically into silicone oil (Clearco, density
 $\rho = 0.970\,\rm g\,cm^{-3}$
, viscosity
$\rho = 0.970\,\rm g\,cm^{-3}$
, viscosity
 $\eta = 105\ \rm Pa\ s$
) contained in a plastic cubic tank of side length
$\eta = 105\ \rm Pa\ s$
) contained in a plastic cubic tank of side length
 $50R$
. To impose a cylindrical boundary, we submerge a thin PET tube of radius
$50R$
. To impose a cylindrical boundary, we submerge a thin PET tube of radius
 $R_0 = 25R$
with a refractive index close to that of silicone oil in the tank. The axes of the two helices define an axial plane (the
$R_0 = 25R$
with a refractive index close to that of silicone oil in the tank. The axes of the two helices define an axial plane (the
 $x$
–
$x$
–
 $z$
plane). The origin ‘O’ of the orthogonal
$z$
plane). The origin ‘O’ of the orthogonal
 $x$
–
$x$
–
 $y$
plane is set at the middle point connecting the axes of the two helices (figure 1
c). The axis of the tube is carefully aligned within the axial plane through the origin. Two separate stepper motors (NEMA 8) drive the rotation of the helices at constant angular speed
$y$
plane is set at the middle point connecting the axes of the two helices (figure 1
c). The axis of the tube is carefully aligned within the axial plane through the origin. Two separate stepper motors (NEMA 8) drive the rotation of the helices at constant angular speed
 $\omega= 0.5 \,\rm rad\,s^{-1}$
in the clockwise direction when looking from above, which sets a velocity scale
$\omega= 0.5 \,\rm rad\,s^{-1}$
in the clockwise direction when looking from above, which sets a velocity scale
 $\omega R= 0.5 \,\rm cm\,s^{-1}$
with Reynolds number
$\omega R= 0.5 \,\rm cm\,s^{-1}$
with Reynolds number
 ${Re} = \rho \omega R^2/\eta \approx 4 \times 10^{-4}$
. The distance
${Re} = \rho \omega R^2/\eta \approx 4 \times 10^{-4}$
. The distance
 $d$
between the axes of the two helices, and their phase difference
$d$
between the axes of the two helices, and their phase difference
 $\Delta \phi$
, are varied in our experiments. Two reaction torque sensors (TFF 400, FUTEK), one for each helix, are mounted on a supporting rail and connected to the stepper motors, which measure the torque applied by the motors to the helices. The supporting rail can rotate on a circular track concentric with the tube axis, enabling us to adjust the orientation of the axial plane
$\Delta \phi$
, are varied in our experiments. Two reaction torque sensors (TFF 400, FUTEK), one for each helix, are mounted on a supporting rail and connected to the stepper motors, which measure the torque applied by the motors to the helices. The supporting rail can rotate on a circular track concentric with the tube axis, enabling us to adjust the orientation of the axial plane
 $\theta$
.
$\theta$
.

Figure 1. Experimental set-up. (a) Image of the two helices in silicone oil seeded with PIV tracers. The helical radius
 $R$
, axial length
$R$
, axial length
 $L$
, pitch length
$L$
, pitch length
 $\lambda$
, filament radius
$\lambda$
, filament radius
 $a$
, and inter-flagellar spacing
$a$
, and inter-flagellar spacing
 $d$
are marked. (b,c) Side and top views of the experimental apparatus. Stepper motors are mounted on a supporting rail attached to two sliding carts, which can rotate along a circular track by an angle
$d$
are marked. (b,c) Side and top views of the experimental apparatus. Stepper motors are mounted on a supporting rail attached to two sliding carts, which can rotate along a circular track by an angle
 $\theta$
. (d) Schematic showing a plane, normal to the axes of two helices, with the phase difference
$\theta$
. (d) Schematic showing a plane, normal to the axes of two helices, with the phase difference
 $\Delta \phi$
. Dashed circles indicate the trajectories of the helices in the plane.
$\Delta \phi$
. Dashed circles indicate the trajectories of the helices in the plane.
2.2. Particle image velocimetry analysis and slender-body theory model
We use particle image velocimetry (PIV) to measure the three-dimensional (3-D) flow field of the rotating helices (PIVLab, Matlab). Silicone oil is seeded with polyamide microparticles (LaVision,
 $\rho = 1.03\,\rm g\,cm^{-3}$
, diameter 60
$\rho = 1.03\,\rm g\,cm^{-3}$
, diameter 60
 $\unicode{x03BC}$
m) at concentration 0.01
$\unicode{x03BC}$
m) at concentration 0.01
 $\%$
(v/v). Two 50 mW, 532 nm continuous line lasers (CivilLaser), positioned on opposite sides of the tank, generate an aligned laser sheet approximately 1 mm thick in the measurement window through the tube axis. By varying the orientation of the axial plane with respect to the laser sheet, we image the axial flow at different polar angles
$\%$
(v/v). Two 50 mW, 532 nm continuous line lasers (CivilLaser), positioned on opposite sides of the tank, generate an aligned laser sheet approximately 1 mm thick in the measurement window through the tube axis. By varying the orientation of the axial plane with respect to the laser sheet, we image the axial flow at different polar angles
 $\theta$
(see figure 1(c) and supplementary movie 1). The flow in the
$\theta$
(see figure 1(c) and supplementary movie 1). The flow in the
 $x$
–
$x$
–
 $y$
plane is captured from the bottom of the tank using a horizontal laser sheet and a 45-degree inclined mirror (supplementary movie 2). Videos are recorded at 5 fps using a CMOS camera (Basler acA2040-90um). The spatial resolution of PIV is 1.3 mm in the
$y$
plane is captured from the bottom of the tank using a horizontal laser sheet and a 45-degree inclined mirror (supplementary movie 2). Videos are recorded at 5 fps using a CMOS camera (Basler acA2040-90um). The spatial resolution of PIV is 1.3 mm in the
 $x$
–
$x$
–
 $y$
plane, and 4.4 mm in the
$y$
plane, and 4.4 mm in the
 $x$
–
$x$
–
 $z$
plane. The root mean square velocity error is
$z$
plane. The root mean square velocity error is
 $0.03 \omega R$
. We also simulate the flow field of two rotating helices using slender-body theory (SBT). The SBT formulas from Rodenborn et al. (Reference Rodenborn, Chen, Swinney, Liu and Zhang2013) for a single helix are extended for two rotating helices with different
$0.03 \omega R$
. We also simulate the flow field of two rotating helices using slender-body theory (SBT). The SBT formulas from Rodenborn et al. (Reference Rodenborn, Chen, Swinney, Liu and Zhang2013) for a single helix are extended for two rotating helices with different
 $d$
and
$d$
and
 $\Delta \phi$
(see Appendix A).
$\Delta \phi$
(see Appendix A).
3. Results
3.1. Torque
We first measure the time-averaged torque required to rotate a single helix in a helical pair, where two helices, separated by distance
 $d$
, rotate at the same speed with a phase difference
$d$
, rotate at the same speed with a phase difference
 $\Delta \phi$
(see figure 2). The torque is normalised by
$\Delta \phi$
(see figure 2). The torque is normalised by
 $T_s$
, the torque required to rotate a single helix at the centre of the tube at the same speed. At large separations,
$T_s$
, the torque required to rotate a single helix at the centre of the tube at the same speed. At large separations,
 $T/T_s \to 1$
, indicating negligible hydrodynamic interactions between distant helices. As the helices move closer,
$T/T_s \to 1$
, indicating negligible hydrodynamic interactions between distant helices. As the helices move closer,
 $T$
displays stronger deviation from
$T$
displays stronger deviation from
 $T_s$
:
$T_s$
:
 $T \lt T_s$
for
$T \lt T_s$
for
 $\Delta \phi \lt \unicode{x03C0} /2$
, whereas
$\Delta \phi \lt \unicode{x03C0} /2$
, whereas
 $T \gt T_s$
for
$T \gt T_s$
for
 $\Delta \phi \gt \unicode{x03C0} /2$
. Interestingly, the hydrodynamic coupling between the helices vanishes at a critical phase difference
$\Delta \phi \gt \unicode{x03C0} /2$
. Interestingly, the hydrodynamic coupling between the helices vanishes at a critical phase difference
 $\Delta \phi = \Delta \phi _c \approx \unicode{x03C0} /2$
, leaving
$\Delta \phi = \Delta \phi _c \approx \unicode{x03C0} /2$
, leaving
 $T = T_s$
regardless of their spacing. The SBT calculation in an unbounded fluid quantitatively matches our experiments (figure 2), except at small
$T = T_s$
regardless of their spacing. The SBT calculation in an unbounded fluid quantitatively matches our experiments (figure 2), except at small
 $d$
, where higher-order near-field interactions ignored in SBT become important due to the close proximity of the helices. The overall agreement suggests that the system boundary at
$d$
, where higher-order near-field interactions ignored in SBT become important due to the close proximity of the helices. The overall agreement suggests that the system boundary at
 $R_0 = 25R$
has a minimal effect.
$R_0 = 25R$
has a minimal effect.

Figure 2. The torque required to rotate one helix in a helical pair,
 $T$
, as a function of the spacing between the helices,
$T$
, as a function of the spacing between the helices,
 $d$
, at different phase differences
$d$
, at different phase differences
 $\Delta \phi$
. Here,
$\Delta \phi$
. Here,
 $T$
is normalised by the torque required to rotate a single helix positioned at the centre of the system,
$T$
is normalised by the torque required to rotate a single helix positioned at the centre of the system,
 $T_s$
, and
$T_s$
, and
 $d$
is normalised by the helical radius
$d$
is normalised by the helical radius
 $R$
. Symbols are experimental data, while the lines show the corresponding SBT calculations.
$R$
. Symbols are experimental data, while the lines show the corresponding SBT calculations.
While the impact of hydrodynamic interactions on the torque of a flagellar bundle has been explored in simulations (Kim & Powers Reference Kim and Powers2004; Buchmann et al. Reference Buchmann, Fauci, Leiderman, Strawbridge and Zhao2018; Tătulea-Codrean & Lauga Reference Tătulea-Codrean and Lauga2021), we provide direct experimental evidence of this non-trivial effect, which has significant implications for the dynamics of bacterial flagellar bundles. In particular, during bundle formation, flagella must synchronise their rotation (
 $\Delta \phi \to 0$
) while reducing their separation (
$\Delta \phi \to 0$
) while reducing their separation (
 $d \to 0$
). As the torque and the rotation speed of a helix are linearly correlated in Stokes flow via a drag coefficient
$d \to 0$
). As the torque and the rotation speed of a helix are linearly correlated in Stokes flow via a drag coefficient
 $\xi$
, i.e.
$\xi$
, i.e.
 $T=\xi \omega$
, our results show that
$T=\xi \omega$
, our results show that
 $\xi$
of a rotating helix in a helical pair decreases with decreasing
$\xi$
of a rotating helix in a helical pair decreases with decreasing
 $d$
and
$d$
and
 $\Delta \phi$
, leading to the observed decrease in torque when the helices rotate at a constant speed (figure 2). For bacterial flagella that operate under constant torque (Berg Reference Berg2004), this decrease in drag coefficient results in an increase in flagellar rotation speed
$\Delta \phi$
, leading to the observed decrease in torque when the helices rotate at a constant speed (figure 2). For bacterial flagella that operate under constant torque (Berg Reference Berg2004), this decrease in drag coefficient results in an increase in flagellar rotation speed
 $\omega =T/\xi$
as
$\omega =T/\xi$
as
 $d\to0$
and
$d\to0$
and
 $\Delta \phi \to 0$
during bundle formation. Notably,
$\Delta \phi \to 0$
during bundle formation. Notably,
 $\xi$
depends only on the geometry of the system and the fluid viscosity, regardless of whether the helices rotate at a constant speed or constant torque. Hence while the power required to rotate a helix,
$\xi$
depends only on the geometry of the system and the fluid viscosity, regardless of whether the helices rotate at a constant speed or constant torque. Hence while the power required to rotate a helix,
 $T\omega = \xi \omega ^2$
, decreases with
$T\omega = \xi \omega ^2$
, decreases with
 $\xi$
as
$\xi$
as
 $d$
and
$d$
and
 $\Delta \phi$
reduce in our case with constant
$\Delta \phi$
reduce in our case with constant
 $\omega$
,
$\omega$
,
 $T\omega = T^2/\xi$
increases with decreasing
$T\omega = T^2/\xi$
increases with decreasing
 $\xi$
for bacteria during bundle formation under constant
$\xi$
for bacteria during bundle formation under constant
 $T$
. Thus unlike in-phase beating of two undulating sheets, which minimises energy dissipation (Taylor Reference Taylor1951), flagellar synchronisation is not a power-saving mechanism for bacteria (Reichert & Stark Reference Reichert and Stark2005; Kanehl & Ishikawa Reference Kanehl and Ishikawa2014). Our results also indicate that increasing the number of flagella in a bundle reduces the drag coefficient of each flagellum in the bundle, allowing flagella to rotate at higher speeds under constant torque (Kamdar et al. Reference Kamdar2023).
$T$
. Thus unlike in-phase beating of two undulating sheets, which minimises energy dissipation (Taylor Reference Taylor1951), flagellar synchronisation is not a power-saving mechanism for bacteria (Reichert & Stark Reference Reichert and Stark2005; Kanehl & Ishikawa Reference Kanehl and Ishikawa2014). Our results also indicate that increasing the number of flagella in a bundle reduces the drag coefficient of each flagellum in the bundle, allowing flagella to rotate at higher speeds under constant torque (Kamdar et al. Reference Kamdar2023).

Figure 3. Hydrodynamic interactions between rotating helices. (a–d) The
 $x$
–
$x$
–
 $y$
flow field of a single rotating helix with its axis positioned at
$y$
flow field of a single rotating helix with its axis positioned at
 $(R,0)$
over one rotation cycle obtained from PIV. White lines indicate streamlines, while the colour represents the magnitude of the in-plane velocity
$(R,0)$
over one rotation cycle obtained from PIV. White lines indicate streamlines, while the colour represents the magnitude of the in-plane velocity
 $v_{x-y}$
, normalised by the rotation speed
$v_{x-y}$
, normalised by the rotation speed
 $\omega R$
(see colour bar below (e)). The solid black line represents the circular trajectory of the real helix, with its cross-section in the plane marked by a black dot. When an imaginary helix is placed at
$\omega R$
(see colour bar below (e)). The solid black line represents the circular trajectory of the real helix, with its cross-section in the plane marked by a black dot. When an imaginary helix is placed at
 $(-2R, 0)$
, it also traces a circular path, shown as the dashed circle. The empty symbols mark the position of the imaginary helix when the phase difference between the real and imaginary helices is
$(-2R, 0)$
, it also traces a circular path, shown as the dashed circle. The empty symbols mark the position of the imaginary helix when the phase difference between the real and imaginary helices is
 $\Delta \phi = 0$
(diamond, left),
$\Delta \phi = 0$
(diamond, left),
 $\Delta\phi=\unicode{x03C0}/2$
(triangle, bottom), and
$\Delta\phi=\unicode{x03C0}/2$
(triangle, bottom), and
 $\Delta\phi=\unicode{x03C0}$
(square, right), respectively. Red arrows indicate the rotation velocity of the real and imaginary helices. (e) The torque on an imaginary helix in the flow field of a real helix,
$\Delta\phi=\unicode{x03C0}$
(square, right), respectively. Red arrows indicate the rotation velocity of the real and imaginary helices. (e) The torque on an imaginary helix in the flow field of a real helix,
 $\langle v_\theta ^r\rangle /\omega R$
, approximates the torque on one helix in the helical pair,
$\langle v_\theta ^r\rangle /\omega R$
, approximates the torque on one helix in the helical pair,
 $T/T_s$
. Symbols are experimental measurements of
$T/T_s$
. Symbols are experimental measurements of
 $T/T_s$
, the same as those in figure 2. Lines represent
$T/T_s$
, the same as those in figure 2. Lines represent
 $\langle v_{\theta }^r \rangle / \omega R$
.
$\langle v_{\theta }^r \rangle / \omega R$
.
3.2. Hydrodynamic interaction
To illustrate the effect of hydrodynamic interactions between two rotating helices on the torque, we first show the PIV flow field of a single rotating helix in the
 $x$
–
$x$
–
 $y$
plane (figure 3
a). Next, we introduce an imaginary helix, placed at distance
$y$
plane (figure 3
a). Next, we introduce an imaginary helix, placed at distance
 $d = 3R$
to the left of the real helix, that experiences but does not disturb the flow of the real helix. This imaginary helix enables a ‘one-way’ hydrodynamic coupling, where the imaginary helix experiences the flow and the resulting hydrodynamic force from the real helix, but does not influence the real helix. Consider three phase differences,
$d = 3R$
to the left of the real helix, that experiences but does not disturb the flow of the real helix. This imaginary helix enables a ‘one-way’ hydrodynamic coupling, where the imaginary helix experiences the flow and the resulting hydrodynamic force from the real helix, but does not influence the real helix. Consider three phase differences,
 $\Delta \phi = 0$
,
$\Delta \phi = 0$
,
 $\unicode{x03C0} /2$
and
$\unicode{x03C0} /2$
and
 $\unicode{x03C0}$
, represented by the three positions of the imaginary helix (three empty symbols along the dashed circle in figure 3
a). At
$\unicode{x03C0}$
, represented by the three positions of the imaginary helix (three empty symbols along the dashed circle in figure 3
a). At
 $\Delta \phi = 0$
(empty diamond on the left), the flow of the real helix aligns with the rotation direction of the imaginary helix. As a result, the imaginary helix rotates at a lower speed relative to the local flow, experiencing reduced fluid drag and consequently smaller torque. In contrast, at
$\Delta \phi = 0$
(empty diamond on the left), the flow of the real helix aligns with the rotation direction of the imaginary helix. As a result, the imaginary helix rotates at a lower speed relative to the local flow, experiencing reduced fluid drag and consequently smaller torque. In contrast, at
 $\Delta \phi = \unicode{x03C0}$
(empty square on the right), the local flow opposes the rotation direction of the imaginary helix. The imaginary helix rotates at a higher speed relative to the local flow, resulting in increased drag and larger torque. Finally, at
$\Delta \phi = \unicode{x03C0}$
(empty square on the right), the local flow opposes the rotation direction of the imaginary helix. The imaginary helix rotates at a higher speed relative to the local flow, resulting in increased drag and larger torque. Finally, at
 $\Delta \phi = \unicode{x03C0} /2$
near
$\Delta \phi = \unicode{x03C0} /2$
near
 $\Delta \phi _c$
(empty triangle at the bottom), the flow is nearly perpendicular to the rotation direction of the imaginary helix, causing minimal modification to the torque experienced by the imaginary helix. As the helices rotate, their relative positions evolve over time (figures 3
a–d). Nevertheless, throughout the rotation cycle, the relative orientation between the rotation of the imaginary helix and the local flow remains qualitatively similar, except when a flow vortex passes near the imaginary helix (figure 3
c). During this phase, however, the magnitude of the local flow drops significantly due to the presence of the vortex, minimising the hydrodynamic interaction between the helices.
$\Delta \phi _c$
(empty triangle at the bottom), the flow is nearly perpendicular to the rotation direction of the imaginary helix, causing minimal modification to the torque experienced by the imaginary helix. As the helices rotate, their relative positions evolve over time (figures 3
a–d). Nevertheless, throughout the rotation cycle, the relative orientation between the rotation of the imaginary helix and the local flow remains qualitatively similar, except when a flow vortex passes near the imaginary helix (figure 3
c). During this phase, however, the magnitude of the local flow drops significantly due to the presence of the vortex, minimising the hydrodynamic interaction between the helices.
Quantitatively, we calculate the velocity of the imaginary helix in the reference frame of the local flow of the single real helix at time
 $t$
,
$t$
,
 $v_{\theta }^r(t) = \omega R - v_l(t)$
, where
$v_{\theta }^r(t) = \omega R - v_l(t)$
, where
 $\omega R$
is the rotation velocity of the imaginary helix in the lab frame, and
$\omega R$
is the rotation velocity of the imaginary helix in the lab frame, and
 $v_l(t)$
is the local flow velocity of the real helix projected onto the rotation direction of the imaginary helix. We then take a time average over one rotation period:
$v_l(t)$
is the local flow velocity of the real helix projected onto the rotation direction of the imaginary helix. We then take a time average over one rotation period:
 $\langle v_{\theta }^r \rangle = (\omega /2\unicode{x03C0} )\int _0^{2\unicode{x03C0} /\omega } v_{\theta }^r(t)\,{\rm d}t$
. Due to the periodicity of the helices, this cycle average at a fixed height
$\langle v_{\theta }^r \rangle = (\omega /2\unicode{x03C0} )\int _0^{2\unicode{x03C0} /\omega } v_{\theta }^r(t)\,{\rm d}t$
. Due to the periodicity of the helices, this cycle average at a fixed height
 $z$
is equivalent to a spatial average over a pitch at a fixed time
$z$
is equivalent to a spatial average over a pitch at a fixed time
 $t$
, apart from minor end effects. As the torque on a helix is proportional to its rotation speed, the torque on the imaginary helix in the reference frame of the local flow of the real helix, relative to the torque of the real helix in the lab frame, is
$t$
, apart from minor end effects. As the torque on a helix is proportional to its rotation speed, the torque on the imaginary helix in the reference frame of the local flow of the real helix, relative to the torque of the real helix in the lab frame, is
 $\langle v_{\theta }^r \rangle / \omega R$
, which serves as a good estimate of the normalised torque on a helix in the helical pair,
$\langle v_{\theta }^r \rangle / \omega R$
, which serves as a good estimate of the normalised torque on a helix in the helical pair,
 $T/T_s$
(figure 3
e).
$T/T_s$
(figure 3
e).
3.3. Confinement

Figure 4. Hydrodynamic interactions between rotating helices under confinement (a–c)
 $R_0 = 7R$
and (d–f)
$R_0 = 7R$
and (d–f)
 $R_0 = 3.5R$
. (a,d) Torque on one helix in the helical pair as a function of phase difference
$R_0 = 3.5R$
. (a,d) Torque on one helix in the helical pair as a function of phase difference
 $\Delta \phi$
at different spacings
$\Delta \phi$
at different spacings
 $d$
. Symbols denote experimental data, while dotted lines represent model predictions (3.2). The green dashed line in (d) is the model prediction using experimentally obtained
$d$
. Symbols denote experimental data, while dotted lines represent model predictions (3.2). The green dashed line in (d) is the model prediction using experimentally obtained
 $T_e/T_c$
, instead of (3.1). (b,e) Flow field in the
$T_e/T_c$
, instead of (3.1). (b,e) Flow field in the
 $x$
–
$x$
–
 $y$
plane of a rotating helix positioned at
$y$
plane of a rotating helix positioned at
 $(R,0)$
. The circle on the right indicates the helix’s trajectory, while the dashed circle on the left represents the trajectory of an imaginary helix. Three phase differences between the real helix and the imaginary helix are shown, consistent with those in figure 3(a). (c,f) Velocity of the imaginary helix in the reference frame of local flow,
$(R,0)$
. The circle on the right indicates the helix’s trajectory, while the dashed circle on the left represents the trajectory of an imaginary helix. Three phase differences between the real helix and the imaginary helix are shown, consistent with those in figure 3(a). (c,f) Velocity of the imaginary helix in the reference frame of local flow,
 $\langle v_\theta ^r \rangle$
, normalised by the helix’s rotation speed in the lab frame,
$\langle v_\theta ^r \rangle$
, normalised by the helix’s rotation speed in the lab frame,
 $\omega R$
, as a function of
$\omega R$
, as a function of
 $\Delta \phi$
at different
$\Delta \phi$
at different
 $d$
.
$d$
.
Bacteria often inhabit confined or structured environments, such as soil pores, intestines and biofilms, where their motility is significantly altered (Vizsnyiczai et al. Reference Vizsnyiczai, Frangipane, Bianchi, Saglimbeni, Dell’Arciprete and Di Leonardo2020). To understand the pumping and mixing of rotating helices in narrow microfluidic channels, it is also important to resolve the dynamics of rotating helices under confinement. However, a SBT formulation for rotating helices in cylindrically confined geometries remains unavailable. Instead, we experimentally investigate a pair of rotating helices under confinement using tubes of varying radii
 $R_0$
, and extend our simple model of hydrodynamic interactions to predict the torque on confined helices.
$R_0$
, and extend our simple model of hydrodynamic interactions to predict the torque on confined helices.
Figures 4(a) and 4(d) show the torque on one helix in the helical pair as a function of phase difference at varying spacings under two different confinement conditions. Similar to the weakly confined system, the torque increases with
 $\Delta \phi$
, and this increasing trend weakens at large spacings. However, under confinement, the mean torque shifts to higher values as
$\Delta \phi$
, and this increasing trend weakens at large spacings. However, under confinement, the mean torque shifts to higher values as
 $d$
increases. Consequently, the critical phase difference
$d$
increases. Consequently, the critical phase difference
 $\Delta \phi _c$
, at which the hydrodynamic interactions disappear, decreases with increasing
$\Delta \phi _c$
, at which the hydrodynamic interactions disappear, decreases with increasing
 $d$
, and eventually vanishes at large
$d$
, and eventually vanishes at large
 $d$
.
$d$
.
Following the method in § 3.2, we estimate the torque on the helices from the flow field of a single helix located near the centre of the confining tube (figure 4
b,e). The cycle-averaged relative velocity of a rotating imaginary helix in the flow field of the single helix
 $\langle v_\theta ^r \rangle$
is computed at different
$\langle v_\theta ^r \rangle$
is computed at different
 $\Delta \phi$
and
$\Delta \phi$
and
 $d$
(figure 4
c,f). While
$d$
(figure 4
c,f). While
 $\langle v_\theta ^r \rangle /\omega R$
captures the trend of
$\langle v_\theta ^r \rangle /\omega R$
captures the trend of
 $T/T_s$
, it fails to predict the upward shift in the mean torque with increasing
$T/T_s$
, it fails to predict the upward shift in the mean torque with increasing
 $d$
. As a result, the predicted
$d$
. As a result, the predicted
 $\Delta \phi _c$
remains close to
$\Delta \phi _c$
remains close to
 $\unicode{x03C0} /2$
regardless of confinement.
$\unicode{x03C0} /2$
regardless of confinement.
The offset of the measured mean torque arises because of the eccentric positioning of the helices and their proximity to the system boundary at large
 $d$
in confined systems. As the effect of the shape of an object on fluid drag is significantly subdued in Stokes flow, we hypothesise that the cycle-averaged torque on a rotating helix within a tube can be approximated by the torque on a rotating cylinder in the tube, where the radius of the cylinder is equal to the helical radius of the helix
$d$
in confined systems. As the effect of the shape of an object on fluid drag is significantly subdued in Stokes flow, we hypothesise that the cycle-averaged torque on a rotating helix within a tube can be approximated by the torque on a rotating cylinder in the tube, where the radius of the cylinder is equal to the helical radius of the helix
 $R$
. The ratio between the torque on an eccentric cylinder
$R$
. The ratio between the torque on an eccentric cylinder
 $T_e$
and that on a concentric cylinder
$T_e$
and that on a concentric cylinder
 $T_c$
is given as (Jeffery Reference Jeffery1922; Yamagata Reference Yamagata1970)
$T_c$
is given as (Jeffery Reference Jeffery1922; Yamagata Reference Yamagata1970)
 \begin{align} &\frac {T_e}{T_c}=\frac {1}{G} \frac {P}{Q} \frac {1}{1+(e / \delta )^2 / 2},\nonumber \\ &\text{with}\quad P=1+2(e / \delta )^2 \frac {1+(\delta / 2 R)-(\delta / 2 R)^2} {(1+\delta / R)^2}-\frac {1}{2} \frac {(e / \delta )^4(\delta / R)^2}{(1+\delta / R)^2}, \nonumber \\ &Q=\left [1-\frac {(e / \delta )^2}{\left [1+\frac {1}{2}(\delta / R)(1-e / \delta )^2\right ]^2}\right ]^{{1}/{2}}\quad \mathrm{and}\quad G=1-\frac {1}{2}(e / \delta )^2 \frac {\delta / R}{1+(\delta / 2 R)}. \end{align}
\begin{align} &\frac {T_e}{T_c}=\frac {1}{G} \frac {P}{Q} \frac {1}{1+(e / \delta )^2 / 2},\nonumber \\ &\text{with}\quad P=1+2(e / \delta )^2 \frac {1+(\delta / 2 R)-(\delta / 2 R)^2} {(1+\delta / R)^2}-\frac {1}{2} \frac {(e / \delta )^4(\delta / R)^2}{(1+\delta / R)^2}, \nonumber \\ &Q=\left [1-\frac {(e / \delta )^2}{\left [1+\frac {1}{2}(\delta / R)(1-e / \delta )^2\right ]^2}\right ]^{{1}/{2}}\quad \mathrm{and}\quad G=1-\frac {1}{2}(e / \delta )^2 \frac {\delta / R}{1+(\delta / 2 R)}. \end{align}
Here,
 $\delta = R_0-R$
is the difference between the radius of the cylinder and that of the tube, and
$\delta = R_0-R$
is the difference between the radius of the cylinder and that of the tube, and
 $e/\delta = d/(2\delta )$
is the eccentricity (figure 5 inset). To verify our hypothesis, we first identify the critical phase difference
$e/\delta = d/(2\delta )$
is the eccentricity (figure 5 inset). To verify our hypothesis, we first identify the critical phase difference
 $\Delta \phi _c$
at each
$\Delta \phi _c$
at each
 $d$
and
$d$
and
 $R_0$
from our calculation of
$R_0$
from our calculation of
 $\langle v_\theta ^r \rangle$
(figure 4
c,f), where the hydrodynamic interactions between the two helices are nullified. We then obtain
$\langle v_\theta ^r \rangle$
(figure 4
c,f), where the hydrodynamic interactions between the two helices are nullified. We then obtain
 $T / T_s$
at
$T / T_s$
at
 $\Delta \phi _c$
from experiments via interpolation (figure 4
a,d), and compare the results with the prediction of (3.1) (figure 5). Experiments and the model exhibit good quantitative agreement in the weakly and moderately confined geometries, supporting our hypothesis. Not surprisingly, (3.1) overestimates the torque by
$\Delta \phi _c$
from experiments via interpolation (figure 4
a,d), and compare the results with the prediction of (3.1) (figure 5). Experiments and the model exhibit good quantitative agreement in the weakly and moderately confined geometries, supporting our hypothesis. Not surprisingly, (3.1) overestimates the torque by
 $\sim 20\%$
for the most confined geometry when the helix is too close to the wall at large
$\sim 20\%$
for the most confined geometry when the helix is too close to the wall at large
 $d$
, where a rotating helix can no longer be approximated accurately by a rotating cylinder.
$d$
, where a rotating helix can no longer be approximated accurately by a rotating cylinder.

Figure 5. Torque on a cylinder or helix
 $T_e$
in tubes of radius
$T_e$
in tubes of radius
 $R_0$
as a function of the eccentricity
$R_0$
as a function of the eccentricity
 $e/\delta$
. Here,
$e/\delta$
. Here,
 $T_e$
is normalised by the torque of a helix rotating concentrically in the tubes,
$T_e$
is normalised by the torque of a helix rotating concentrically in the tubes,
 $T_c$
. Symbols are from experiments, and lines are from (3.1). Inset: schematic illustrating the position of the cylinder/helix within a tube.
$T_c$
. Symbols are from experiments, and lines are from (3.1). Inset: schematic illustrating the position of the cylinder/helix within a tube.
The quantitative agreement shown in figure 5 suggests that we can decompose the effect of the confining boundary on the torque of rotating helices into two parts. (i) The boundary modifies the flow field of the helices, altering their mutual hydrodynamic interactions, as reflected in the trend of
 $\langle v_\theta ^r\rangle /\omega R$
. (ii) The helices experience additional drag,
$\langle v_\theta ^r\rangle /\omega R$
. (ii) The helices experience additional drag,
 $(T_e-T_c)/T_c$
, from the hydrodynamic interactions with the boundary due to their eccentric positions within the confining tube. This decomposition of the confinement-induced hydrodynamic interactions into pair-induced and self-induced contributions is similar to the approach adopted in a recent study on hydrodynamic bound states of rotating cylinders under confinement (Guo, Man & Zhu Reference Guo, Man and Zhu2024). By summing the two contributions,
$(T_e-T_c)/T_c$
, from the hydrodynamic interactions with the boundary due to their eccentric positions within the confining tube. This decomposition of the confinement-induced hydrodynamic interactions into pair-induced and self-induced contributions is similar to the approach adopted in a recent study on hydrodynamic bound states of rotating cylinders under confinement (Guo, Man & Zhu Reference Guo, Man and Zhu2024). By summing the two contributions,
 \begin{equation} \frac {T}{T_s} = \frac {\langle v_\theta ^r\rangle }{\omega R} + \frac {T_e-T_c}{T_c}, \end{equation}
\begin{equation} \frac {T}{T_s} = \frac {\langle v_\theta ^r\rangle }{\omega R} + \frac {T_e-T_c}{T_c}, \end{equation}
we quantitatively predict the torque on a pair of rotating helices across different spacings, phase differences and confinement levels (figures 3(e), 4(a) and 4(d)).
What are the implications of our findings on bacteria swimming under confinement? First, experiments on E. coli show that bacteria tend to swim near solid boundaries in channels of large radii, but shift to swimming along the central axis of more confined channels when the confining radius
 $R_0$
is less than
$R_0$
is less than
 $\sim 7R$
(Vizsnyiczai et al. Reference Vizsnyiczai, Frangipane, Bianchi, Saglimbeni, Dell’Arciprete and Di Leonardo2020). Our results suggest that this axial swimming behaviour, characterised by low eccentricity, reduces the drag coefficient of the flagellar bundle
$\sim 7R$
(Vizsnyiczai et al. Reference Vizsnyiczai, Frangipane, Bianchi, Saglimbeni, Dell’Arciprete and Di Leonardo2020). Our results suggest that this axial swimming behaviour, characterised by low eccentricity, reduces the drag coefficient of the flagellar bundle
 $\xi$
in the torque–speed relation
$\xi$
in the torque–speed relation
 $T=\xi \omega$
(figure 4
a,d). Thus in confined environments, a bacterium is situated at a location where its flagella rotate most rapidly under constant motor torque. Second, our findings reveal that the change of
$T=\xi \omega$
(figure 4
a,d). Thus in confined environments, a bacterium is situated at a location where its flagella rotate most rapidly under constant motor torque. Second, our findings reveal that the change of
 $\xi (d)$
at
$\xi (d)$
at
 $\Delta \phi = 0$
becomes more pronounced in confined systems. This implies that the speed-up of flagellar rotation during bundle formation discussed above should be more evident under confinement – an intriguing prediction that is worth testing experimentally. Finally, although our results support prior predictions that
$\Delta \phi = 0$
becomes more pronounced in confined systems. This implies that the speed-up of flagellar rotation during bundle formation discussed above should be more evident under confinement – an intriguing prediction that is worth testing experimentally. Finally, although our results support prior predictions that
 $\xi$
increases with the degree of confinement (Vizsnyiczai et al. Reference Vizsnyiczai, Frangipane, Bianchi, Saglimbeni, Dell’Arciprete and Di Leonardo2020), our data do not clarify whether confinement enhances or reduces the thrust force of a flagellar bundle (Liu, Breuer & Powers Reference Liu, Breuer and Powers2014). For a translationally fixed rotating helix subjected to a constant torque, the resulting thrust force is
$\xi$
increases with the degree of confinement (Vizsnyiczai et al. Reference Vizsnyiczai, Frangipane, Bianchi, Saglimbeni, Dell’Arciprete and Di Leonardo2020), our data do not clarify whether confinement enhances or reduces the thrust force of a flagellar bundle (Liu, Breuer & Powers Reference Liu, Breuer and Powers2014). For a translationally fixed rotating helix subjected to a constant torque, the resulting thrust force is
 $F = cT/\xi$
, where
$F = cT/\xi$
, where
 $c$
is the off-diagonal term in the drag coefficient matrix that couples the thrust and rotation of the helix,
$c$
is the off-diagonal term in the drag coefficient matrix that couples the thrust and rotation of the helix,
 $F = c\omega$
(Kamdar et al. Reference Kamdar, Shin, Leishangthem, Francis, Xu and Cheng2022, Reference Kamdar2023). While our study shows that
$F = c\omega$
(Kamdar et al. Reference Kamdar, Shin, Leishangthem, Francis, Xu and Cheng2022, Reference Kamdar2023). While our study shows that
 $\xi$
increases with confinement,
$\xi$
increases with confinement,
 $c$
also increases concurrently (Vizsnyiczai et al. Reference Vizsnyiczai, Frangipane, Bianchi, Saglimbeni, Dell’Arciprete and Di Leonardo2020). Future work should aim to directly measure the thrust force of a helical pair under confinement, enabling an estimate of
$c$
also increases concurrently (Vizsnyiczai et al. Reference Vizsnyiczai, Frangipane, Bianchi, Saglimbeni, Dell’Arciprete and Di Leonardo2020). Future work should aim to directly measure the thrust force of a helical pair under confinement, enabling an estimate of
 $c(R_0)$
.
$c(R_0)$
.

Figure 6. The 3-D axial flow field from experiments. (a) Instantaneous
 $v_z(r,z)$
when the cross-sections of the helices at the
$v_z(r,z)$
when the cross-sections of the helices at the
 $z=0$
plane are oriented at
$z=0$
plane are oriented at
 $\phi _1 = \phi _2 =1.28\unicode{x03C0}$
. (b) Time-averaged axial flow field
$\phi _1 = \phi _2 =1.28\unicode{x03C0}$
. (b) Time-averaged axial flow field
 $\langle v_z(r,z)\rangle$
over multiple rotation cycles for
$\langle v_z(r,z)\rangle$
over multiple rotation cycles for
 $\Delta \phi = 0$
and
$\Delta \phi = 0$
and
 $d/R = 6$
in the axial plane (
$d/R = 6$
in the axial plane (
 $\theta = 0$
). (c) Stack of seven time-averaged axial flow fields at different
$\theta = 0$
). (c) Stack of seven time-averaged axial flow fields at different
 $\theta$
. The 3-D axial flow field with (d)
$\theta$
. The 3-D axial flow field with (d)
 $d/R = 2$
and
$d/R = 2$
and
 $\Delta \phi = 0$
, (e)
$\Delta \phi = 0$
, (e)
 $d/R = 2$
and
$d/R = 2$
and
 $\Delta \phi$
=
$\Delta \phi$
=
 $3\unicode{x03C0} /4$
, (f)
$3\unicode{x03C0} /4$
, (f)
 $d/R = 10$
and
$d/R = 10$
and
 $\Delta \phi = 0$
, and(g)
$\Delta \phi = 0$
, and(g)
 $d/R = 10$
and
$d/R = 10$
and
 $\Delta \phi$
=
$\Delta \phi$
=
 $\unicode{x03C0}$
. Dashed circles indicate
$\unicode{x03C0}$
. Dashed circles indicate
 $r_{c}(\theta )$
where
$r_{c}(\theta )$
where
 $\langle v_z \rangle =0$
.
$\langle v_z \rangle =0$
.
3.4. Axial flux and pump efficiency
Finally, we investigate the fluid transport capacity of two rotating helices in the weakly confined system (
 $R_0=25R$
) by imaging the 3-D axial flow. While the radial flow
$R_0=25R$
) by imaging the 3-D axial flow. While the radial flow
 $v_r$
in the axial plane quantitatively agrees with our SBT calculation and previous numerical predictions (Kim & Powers Reference Kim and Powers2004; Buchmann et al. Reference Buchmann, Fauci, Leiderman, Strawbridge and Zhao2018) (see supplementary movie 2), the axial flow
$v_r$
in the axial plane quantitatively agrees with our SBT calculation and previous numerical predictions (Kim & Powers Reference Kim and Powers2004; Buchmann et al. Reference Buchmann, Fauci, Leiderman, Strawbridge and Zhao2018) (see supplementary movie 2), the axial flow
 $v_z$
exhibits qualitatively different behaviour (supplementary movie 1). Figures 6(a) and 6(b) show instantaneous and time-averaged flow fields of the axial flow through the axes of the helices, which reveal a downward flow
$v_z$
exhibits qualitatively different behaviour (supplementary movie 1). Figures 6(a) and 6(b) show instantaneous and time-averaged flow fields of the axial flow through the axes of the helices, which reveal a downward flow
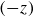 $(-z)$
near the helices, and an upward flow
$(-z)$
near the helices, and an upward flow
 $(+z)$
near the boundary. Unlike in an unbounded fluid, where
$(+z)$
near the boundary. Unlike in an unbounded fluid, where
 $v_z$
decays to zero as
$v_z$
decays to zero as
 $r\to \infty$
,
$r\to \infty$
,
 $v_z$
reaches zero at a finite radius
$v_z$
reaches zero at a finite radius
 $r_c$
, and becomes positive near the wall. The confinement inherent in any experimental system ensures that the net flux across any cross-section of the system normal to the axes of the helices is zero. Due to the lack of axisymmetry in the flow field, the locus
$r_c$
, and becomes positive near the wall. The confinement inherent in any experimental system ensures that the net flux across any cross-section of the system normal to the axes of the helices is zero. Due to the lack of axisymmetry in the flow field, the locus
 $r_c(\theta )$
is non-circular and varies with the azimuthal angle
$r_c(\theta )$
is non-circular and varies with the azimuthal angle
 $\theta$
. We determine
$\theta$
. We determine
 $r_c(\theta )$
for each experiment by interpolating the time-averaged flow field to identify the contour where
$r_c(\theta )$
for each experiment by interpolating the time-averaged flow field to identify the contour where
 $\langle v_z \rangle = 0$
.
$\langle v_z \rangle = 0$
.
To image the 3-D flow field, we rotate the supporting rail and change the polar angle
 $\theta \in [0, \unicode{x03C0} /2]$
(figures 1(c) and 6(c)), leveraging the bilateral symmetry of the system. Representative time-averaged 3-D axial flow fields
$\theta \in [0, \unicode{x03C0} /2]$
(figures 1(c) and 6(c)), leveraging the bilateral symmetry of the system. Representative time-averaged 3-D axial flow fields
 $\langle v_z(r,\theta ) \rangle$
for helices of two phase differences and spacings are shown in figures 6(d)–6(g). We compute the axial flux
$\langle v_z(r,\theta ) \rangle$
for helices of two phase differences and spacings are shown in figures 6(d)–6(g). We compute the axial flux
 $Q = \int _0^{2\unicode{x03C0} } \int _0^{r_c(\theta )} \langle v_z(r, \theta )\rangle r\,{\rm d}r\, {\rm d} \theta$
from these flow fields (figure 7
a). Compared with the flux of two independently rotating helices
$Q = \int _0^{2\unicode{x03C0} } \int _0^{r_c(\theta )} \langle v_z(r, \theta )\rangle r\,{\rm d}r\, {\rm d} \theta$
from these flow fields (figure 7
a). Compared with the flux of two independently rotating helices
 $2Q_s$
, helices pump less fluid when acting in a bundle, regardless of their phase difference
$2Q_s$
, helices pump less fluid when acting in a bundle, regardless of their phase difference
 $\Delta \phi$
or spacing
$\Delta \phi$
or spacing
 $d$
. Similar to torque,
$d$
. Similar to torque,
 $Q$
increases
$Q$
increases
 $\Delta \phi$
, with the increasing trend diminishing at large
$\Delta \phi$
, with the increasing trend diminishing at large
 $d$
. For two helices at the smallest spacing in our experiments,
$d$
. For two helices at the smallest spacing in our experiments,
 $Q \approx 1.2Q_s$
when
$Q \approx 1.2Q_s$
when
 $\Delta \phi = 0$
, indicating that the two closely spaced helices behave similarly to a single rotating helix. In an unbounded fluid, SBT predicts a diverging flux due to the slow decay of the axial flow following
$\Delta \phi = 0$
, indicating that the two closely spaced helices behave similarly to a single rotating helix. In an unbounded fluid, SBT predicts a diverging flux due to the slow decay of the axial flow following
 $\sim 1/r$
at large distances. However, the ratio of the flux of two helices to that of a single helix remains finite, as the pair effectively behaves like a single helix at large distances. Interestingly, despite the qualitative difference in the flow field,
$\sim 1/r$
at large distances. However, the ratio of the flux of two helices to that of a single helix remains finite, as the pair effectively behaves like a single helix at large distances. Interestingly, despite the qualitative difference in the flow field,
 $Q/2Q_s$
from SBT provides a good match to experimental results (figure 7
a).
$Q/2Q_s$
from SBT provides a good match to experimental results (figure 7
a).

Figure 7. Fluid transport and pump efficiency. (a) Normalised axial flux of rotating helices as a function of phase difference
 $\Delta \phi$
for various spacings
$\Delta \phi$
for various spacings
 $d$
. (b) Pump efficiency
$d$
. (b) Pump efficiency
 $\epsilon$
as a function of
$\epsilon$
as a function of
 $\Delta \phi$
for different
$\Delta \phi$
for different
 $d$
. Symbols represent experimental data, while lines correspond to SBT calculations.
$d$
. Symbols represent experimental data, while lines correspond to SBT calculations.
Although helices in a bundle pump less fluid, they also require less power to rotate. We quantify this trade-off by measuring the pump efficiency
 $\epsilon = [Q/(2T\omega )]/[Q_s/(T_s\omega )]$
(Barrett et al. Reference Barrett, Fogelson, Forest, Gruninger, Lim and Griffith2024). Figure 7(b) shows
$\epsilon = [Q/(2T\omega )]/[Q_s/(T_s\omega )]$
(Barrett et al. Reference Barrett, Fogelson, Forest, Gruninger, Lim and Griffith2024). Figure 7(b) shows
 $\epsilon$
for a pair of helices at different
$\epsilon$
for a pair of helices at different
 $\Delta \phi$
and
$\Delta \phi$
and
 $d$
. At small spacings, the pair are more efficient when rotating in phase. However, their efficiency always remains lower than that of two independently rotating helices. The SBT calculation provides a reasonable estimate of the experimental results.
$d$
. At small spacings, the pair are more efficient when rotating in phase. However, their efficiency always remains lower than that of two independently rotating helices. The SBT calculation provides a reasonable estimate of the experimental results.
A swimmer and a pump represent dual manifestations of the same low-
 $Re$
fluid phenomenon: a tethered swimmer functions as a pump, while an unanchored pump behaves as a swimmer (Raz & Avron Reference Raz and Avron2007). Specifically, the fluid flux generated by fixed rotating helices scales linearly with their swimming speed when untethered, while the power required for rotation as a pump is linearly correlated with the power needed for swimming. Thus our results suggest that a bundle of helical filaments is less efficient for swimming than individual filaments rotating at the same speed.
$Re$
fluid phenomenon: a tethered swimmer functions as a pump, while an unanchored pump behaves as a swimmer (Raz & Avron Reference Raz and Avron2007). Specifically, the fluid flux generated by fixed rotating helices scales linearly with their swimming speed when untethered, while the power required for rotation as a pump is linearly correlated with the power needed for swimming. Thus our results suggest that a bundle of helical filaments is less efficient for swimming than individual filaments rotating at the same speed.
4. Conclusion
By integrating experiments with hydrodynamic modelling, we present a study on the dynamics of rotating helices in low-Reynolds-number flow – a fluid phenomenon with fundamental implications for the function of bacterial flagellar bundles, and practical relevance for microfluidics. In particular, we construct a scaled experimental model of a flagellar bundle of two helices, and examine how helix spacing and phase difference influence torque, flow fields and fluid transport. Building upon the flow characteristics of a single helix, we develop a simple model that quantitatively predicts the torque of a pair of rotating helices. We further extend this model to the complex yet important case of rotating helices under confinement, enabling torque calculations that are otherwise inaccessible through existing analytical theories. Finally, we explore the fluid transport capabilities of rotating helices, and assess their efficiency as low-Reynolds-number fluid pumps. Our findings provide key insights into the dynamics of bacterial flagellar bundles and offer valuable guidelines for designing microfluidic pumps.
Supplementary movies
Supplementary movies are available at https://doi.org/10.1017/jfm.2025.10240.
Acknowledgements
We thank D. Retic and A. Dosch for their help with the experimental set-up, and M. Tătulea-Codrean, D. Ghosh, S. Lim and E. Lauga for fruitful discussions.
Funding
The research was supported by NSF CBET 2415405 and University of Minnesota MRSEC.
Declaration of interests
The authors report no conflict of interest.
Appendix A. Slender-body theory
Slender-body theory (SBT) provides a widely used mathematical framework for analysing the dynamics of flagella and cilia in microorganism locomotion (Lauga Reference Lauga2020). We build on the SBT formulation developed by Rodenborn et al. (Reference Rodenborn, Chen, Swinney, Liu and Zhang2013), which relates the force density along a rigid slender helix to its geometry, as well as its rotational and translational velocities in an unbounded fluid. Here, we extend their model to include a second, rotating helix that is parallel to the original and separated from it by distance
 $d$
. The choice of SBT is supported by the work of Martindale, Jabbarzadeh & Fu (Reference Martindale, Jabbarzadeh and Fu2016), which compared several computational methods for modelling helical filaments. The study shows that SBT is the optimal choice, offering a balance of accuracy and computational efficiency for the parameter regime relevant to our system, with
$d$
. The choice of SBT is supported by the work of Martindale, Jabbarzadeh & Fu (Reference Martindale, Jabbarzadeh and Fu2016), which compared several computational methods for modelling helical filaments. The study shows that SBT is the optimal choice, offering a balance of accuracy and computational efficiency for the parameter regime relevant to our system, with
 $\lambda /R = 12$
and
$\lambda /R = 12$
and
 $a/R = 0.1$
. Here,
$a/R = 0.1$
. Here,
 $\lambda$
and
$\lambda$
and
 $R$
are the pitch length and radius of the helix, respectively, and
$R$
are the pitch length and radius of the helix, respectively, and
 $a$
is the radius of the filaments.
$a$
is the radius of the filaments.
We discretise each helix by a set of nodal points
 $\{\varphi _n\}$
along the helical angle
$\{\varphi _n\}$
along the helical angle
 $\varphi (s) = (2\unicode{x03C0} /\lambda )s\cos \theta$
, where
$\varphi (s) = (2\unicode{x03C0} /\lambda )s\cos \theta$
, where
 $\theta = \text{atan}(\lambda /2\unicode{x03C0} R)$
is the pitch angle, and
$\theta = \text{atan}(\lambda /2\unicode{x03C0} R)$
is the pitch angle, and
 $s$
is the arc length. Each helix is approximated by
$s$
is the arc length. Each helix is approximated by
 $N$
straight segments of length
$N$
straight segments of length
 $\Delta s = R\,\Delta \varphi \csc \theta$
, where
$\Delta s = R\,\Delta \varphi \csc \theta$
, where
 $\Delta \varphi$
is the mesh size in the helical phase. Following the parameters used by Rodenborn et al. (Reference Rodenborn, Chen, Swinney, Liu and Zhang2013), we set
$\Delta \varphi$
is the mesh size in the helical phase. Following the parameters used by Rodenborn et al. (Reference Rodenborn, Chen, Swinney, Liu and Zhang2013), we set
 $N = 54$
, yielding a helix of finite length
$N = 54$
, yielding a helix of finite length
 $L = 32R$
that matches the geometry of the helices in our experiments.
$L = 32R$
that matches the geometry of the helices in our experiments.
The centreline of helix 1
 $(n=1,\ldots ,N)$
is
$(n=1,\ldots ,N)$
is
 \begin{equation} \mathbf{r}_1(\varphi _n) = R\left (\cos \varphi _n -\frac {d}{2R},-\sin \varphi _n,\varphi _n\cot \theta \right ), \end{equation}
\begin{equation} \mathbf{r}_1(\varphi _n) = R\left (\cos \varphi _n -\frac {d}{2R},-\sin \varphi _n,\varphi _n\cot \theta \right ), \end{equation}
whereas that of the helix 2
 $(n=N+1,\ldots ,2N)$
is
$(n=N+1,\ldots ,2N)$
is
 \begin{equation} \mathbf{r}_2(\varphi _n) = R\left (\cos \varphi _n + \frac {d}{2R},-\sin \varphi _n,\varphi _n\,\cot \theta \right ). \end{equation}
\begin{equation} \mathbf{r}_2(\varphi _n) = R\left (\cos \varphi _n + \frac {d}{2R},-\sin \varphi _n,\varphi _n\,\cot \theta \right ). \end{equation}
Using the formulation from (18) of Rodenborn et al. (Reference Rodenborn, Chen, Swinney, Liu and Zhang2013), extended to include the second helix with
 $n=1,\ldots ,2N$
, the velocity at node
$n=1,\ldots ,2N$
, the velocity at node
 $n$
is
$n$
is
 \begin{equation} \mathbf{u}_n = \frac {\bigl [\mathbf{I} - \hat {\mathbf{t}}_n\hat {\mathbf{t}}_n^{\rm T} + \mathbf{K}_n\bigr ]\boldsymbol{\cdot} \mathbf{f}_n}{4\unicode{x03C0} \eta } + \frac {\Delta s}{8\unicode{x03C0} \eta } \sum _{\substack {m=1 \\ m\neq n}}^{2N} \left (\frac {\mathbf{I}}{\lvert \mathbf{r}_{nm} \rvert } + \frac {\mathbf{r}_{nm}\mathbf{r}_{nm}^{\rm T}}{\lvert \mathbf{r}_{nm} \rvert ^3}\right) \boldsymbol{\cdot} \mathbf{f}_m, \end{equation}
\begin{equation} \mathbf{u}_n = \frac {\bigl [\mathbf{I} - \hat {\mathbf{t}}_n\hat {\mathbf{t}}_n^{\rm T} + \mathbf{K}_n\bigr ]\boldsymbol{\cdot} \mathbf{f}_n}{4\unicode{x03C0} \eta } + \frac {\Delta s}{8\unicode{x03C0} \eta } \sum _{\substack {m=1 \\ m\neq n}}^{2N} \left (\frac {\mathbf{I}}{\lvert \mathbf{r}_{nm} \rvert } + \frac {\mathbf{r}_{nm}\mathbf{r}_{nm}^{\rm T}}{\lvert \mathbf{r}_{nm} \rvert ^3}\right) \boldsymbol{\cdot} \mathbf{f}_m, \end{equation}
where
 $\mathbf{r}_n=\mathbf{r}_{k(n)}(\varphi _n)$
is the centreline position of node
$\mathbf{r}_n=\mathbf{r}_{k(n)}(\varphi _n)$
is the centreline position of node
 $n$
,
$n$
,
 $\mathbf{r}_{nm} = \mathbf{r}_{k(n)}(\varphi _n) - \mathbf{r}_{k(m)}(\varphi _m)$
is the relative position between two segments
$\mathbf{r}_{nm} = \mathbf{r}_{k(n)}(\varphi _n) - \mathbf{r}_{k(m)}(\varphi _m)$
is the relative position between two segments
 $n$
and
$n$
and
 $m$
, and
$m$
, and
 $k(n) = 1$
or 2 indicates whether segment
$k(n) = 1$
or 2 indicates whether segment
 $n$
belongs to helix 1 or helix 2. The first term denotes local contribution, where
$n$
belongs to helix 1 or helix 2. The first term denotes local contribution, where
 $\hat {\mathbf{t}}_n$
is the local tangent at node
$\hat {\mathbf{t}}_n$
is the local tangent at node
 $n$
,
$n$
,
 $\mathbf{f}_n$
is the unknown force density, and
$\mathbf{f}_n$
is the unknown force density, and
 $\mathbf{K}_n=\ln ({\Delta s}/{2\delta ^{\prime }})\bigl (\mathbf{I}+ \hat {\mathbf{t}}_n\hat {\mathbf{t}}_n^{\rm T}\bigr )$
arises from an asymptotic integral over a small neighbourhood of node
$\mathbf{K}_n=\ln ({\Delta s}/{2\delta ^{\prime }})\bigl (\mathbf{I}+ \hat {\mathbf{t}}_n\hat {\mathbf{t}}_n^{\rm T}\bigr )$
arises from an asymptotic integral over a small neighbourhood of node
 $n$
between the ‘natural cut-off’
$n$
between the ‘natural cut-off’
 $\delta ^{\prime } = a\sqrt e/2$
and
$\delta ^{\prime } = a\sqrt e/2$
and
 $\Delta s/2$
(Lighthill Reference Lighthill1976). Here,
$\Delta s/2$
(Lighthill Reference Lighthill1976). Here,
 $e$
is Euler’s number. The second term accounts for the far-field contribution of all other segments
$e$
is Euler’s number. The second term accounts for the far-field contribution of all other segments
 $m\neq n$
, including those on the other helix. Equation (A3) can be expressed more compactly as a linear mapping:
$m\neq n$
, including those on the other helix. Equation (A3) can be expressed more compactly as a linear mapping:
 \begin{equation} \mathbf{u}^{\rm T} = \mathbf{G}\mathbf{f}^{\rm T}, \end{equation}
\begin{equation} \mathbf{u}^{\rm T} = \mathbf{G}\mathbf{f}^{\rm T}, \end{equation}
where
 $\mathbf{u} = (\mathbf{u}_1,\mathbf{u}_2, \ldots, \mathbf{u}_{2N})$
,
$\mathbf{u} = (\mathbf{u}_1,\mathbf{u}_2, \ldots, \mathbf{u}_{2N})$
,
 $\mathbf{f} = (\mathbf{f}_1,\mathbf{f}_2, \ldots, \mathbf{f}_{2N})$
, and
$\mathbf{f} = (\mathbf{f}_1,\mathbf{f}_2, \ldots, \mathbf{f}_{2N})$
, and
 $\mathbf{G}$
is a
$\mathbf{G}$
is a
 $6N \times 6N$
matrix that captures both self and mutual hydrodynamic interactions. The diagonal terms account for each segment’s self-interaction via the local dipole correction, while the off-diagonal terms represent interactions between all segment pairs
$6N \times 6N$
matrix that captures both self and mutual hydrodynamic interactions. The diagonal terms account for each segment’s self-interaction via the local dipole correction, while the off-diagonal terms represent interactions between all segment pairs
 $(n, m)$
, whether on the same helix or across different helices.
$(n, m)$
, whether on the same helix or across different helices.
For a prescribed motion – such as rigid-body rotation about a fixed axis with angular velocity
 $\mathbf{\Omega }$
, combined with a translation velocity
$\mathbf{\Omega }$
, combined with a translation velocity
 $\mathbf{V}$
– the nodal velocity
$\mathbf{V}$
– the nodal velocity
 $\mathbf{u}_n$
is determined by the no-slip boundary condition
$\mathbf{u}_n$
is determined by the no-slip boundary condition
 \begin{equation} \mathbf{u}_n = \mathbf{\Omega } \times \bigl (\mathbf{r}_n-\mathbf{r}_{{axis}(n)}\bigr )+ \mathbf{V}, \end{equation}
\begin{equation} \mathbf{u}_n = \mathbf{\Omega } \times \bigl (\mathbf{r}_n-\mathbf{r}_{{axis}(n)}\bigr )+ \mathbf{V}, \end{equation}
where
 $\mathbf{r}_{{axis}(n)}$
is the centre of rotation of the helix to which segment
$\mathbf{r}_{{axis}(n)}$
is the centre of rotation of the helix to which segment
 $n$
belongs. By equating (A4) and (A5), we solve for the force densities
$n$
belongs. By equating (A4) and (A5), we solve for the force densities
 $\{\mathbf{f}_n\}_{n=1,\ldots, 2N}$
. The axial force
$\{\mathbf{f}_n\}_{n=1,\ldots, 2N}$
. The axial force
 $F_z$
and torque
$F_z$
and torque
 $T$
on each helix are obtained by summing over the corresponding subset of nodes.
$T$
on each helix are obtained by summing over the corresponding subset of nodes.
Finally, the flow field
 $\mathbf{v(x)}$
is evaluated from
$\mathbf{v(x)}$
is evaluated from
 $\{\mathbf{f}_n\}$
by (Kim & Powers Reference Kim and Powers2004)
$\{\mathbf{f}_n\}$
by (Kim & Powers Reference Kim and Powers2004)
 \begin{equation} v_i(\mathbf{x})=\sum _{n=1}^{2 N}\left [S_{i j}\left (\mathbf{x}, \mathbf{r}_n\right ) \frac {f_{n j}}{8 \unicode{x03C0} \eta }-D_{i j}\left (\mathbf{x}, \mathbf{r}_n\right ) \frac {a^2 f_{n, \perp j}}{16 \unicode{x03C0} \eta }\right ] \Delta s_n, \end{equation}
\begin{equation} v_i(\mathbf{x})=\sum _{n=1}^{2 N}\left [S_{i j}\left (\mathbf{x}, \mathbf{r}_n\right ) \frac {f_{n j}}{8 \unicode{x03C0} \eta }-D_{i j}\left (\mathbf{x}, \mathbf{r}_n\right ) \frac {a^2 f_{n, \perp j}}{16 \unicode{x03C0} \eta }\right ] \Delta s_n, \end{equation}
where
 $i$
and
$i$
and
 $j$
run over
$j$
run over
 $x$
,
$x$
,
 $y$
and
$y$
and
 $z$
. The summation runs over the arc length of both helices,
$z$
. The summation runs over the arc length of both helices,
 $\mathbf{f}_n$
is the force per unit length exerted on the fluid by the
$\mathbf{f}_n$
is the force per unit length exerted on the fluid by the
 $n$
th segment, and
$n$
th segment, and
 $\mathbf{f}_{n \perp }$
is the transverse part of
$\mathbf{f}_{n \perp }$
is the transverse part of
 $\mathbf{f}_n$
. The Stokeslet tensor is
$\mathbf{f}_n$
. The Stokeslet tensor is
 \begin{equation} S_{i j}\left (\mathbf{x}, \mathbf{r}_n\right )=\frac {\delta _{i j}}{\left |\mathbf{x}-\mathbf{r}_n\right |}+\frac {\left (x_i-r_{n, i}\right )\left (x_j-r_{n, j}\right )}{\left |\mathbf{x}-\mathbf{r}_n\right |^3}, \end{equation}
\begin{equation} S_{i j}\left (\mathbf{x}, \mathbf{r}_n\right )=\frac {\delta _{i j}}{\left |\mathbf{x}-\mathbf{r}_n\right |}+\frac {\left (x_i-r_{n, i}\right )\left (x_j-r_{n, j}\right )}{\left |\mathbf{x}-\mathbf{r}_n\right |^3}, \end{equation}
and the doublet tensor is
 \begin{equation} D_{i j}\left (\mathbf{x}, \mathbf{r}_n\right )=-\frac {\delta _{i j}}{\left |\mathbf{x}-\mathbf{r}_n\right |^3}+3 \frac {\left (x_i-r_{n, i}\right )\left (x_j-r_{n, j}\right )}{\left |\mathbf{x}-\mathbf{r}_n\right |^5}. \end{equation}
\begin{equation} D_{i j}\left (\mathbf{x}, \mathbf{r}_n\right )=-\frac {\delta _{i j}}{\left |\mathbf{x}-\mathbf{r}_n\right |^3}+3 \frac {\left (x_i-r_{n, i}\right )\left (x_j-r_{n, j}\right )}{\left |\mathbf{x}-\mathbf{r}_n\right |^5}. \end{equation}















































































 Open access
Open access