


Introduction
Wolf poison or bloody-root stellera (Stellera chamaejasme L.) belongs to the family Thymelaeaceae. It is a perennial herbaceous plant (Zhang et al. Reference Zhang, Volis and Sun2010). According to the Flora of China (Editorial Committee of the Flora of China, 2004), S. chamaejasme thrives in harsh environments often found at higher altitudes on dry, sunny slopes of mountains and alpine grasslands. The plant forms dense clumps, produces nut-like fruits, and has a long life span (Li et al. Reference Li, Xiang, Tang, Jiang, Duang and Chang2019). Traditionally, S. chamaejasme has been used in Chinese medicine. Its roots are employed for their bitter and neutral properties, believed to have effects such as antimicrobial, insecticidal, dispersing, diuretic, analgesic, and expectorant properties (L Wu et al. Reference Wu, Pu, Gao, Chen, Tao, Liu and Hou2014; Zhang et al. Reference Zhang, Gao, Zhang, Zhang, Wang and Sun2016). However, its marked toxicity induces acute poisoning in grazing animals, potentially leading to fatalities, with specific components, notably the stelleraterpenoids A-M class, exhibiting significant cytotoxic activity (Pan et al. Reference Pan, Su, Liu, Deng, Hu, Wu, Xia, Chen, He, Chen and Wan2021; You et al. Reference You, Ma, Guo, Kong, Shi, Wu and Zhao2018). Modern pharmacological studies have further revealed significant pharmacological activities associated with S. chamaejasme, including antibacterial and anti-inflammatory effects (Kim et al. Reference Kim, Lee, Randy, Yun, Oh and Nho2017; Selenge et al. Reference Selenge, Vieira, Gendaram, Reis, Tsolmon, Tsendeekhuu, Ferreira and Neves2023), antiviral properties (Asada et al. Reference Asada, Sukemori, Watanabe, Malla, Yoshikawa, Li, Koike, Chen, Akiyama, Qian, Nakagawa-Goto, Morris-Natschke and Lee2011; Shahrajabian and Sun, Reference Shahrajabian and Sun2023), antitumor activity (Li et al. Reference Li, Wang, Xiao, Li, Kan, Wang, Zhang, Wang, Yang, Chen, Weng, Chen, Zhou, Guo, Liu and Zhu2016; Yu et al. Reference Yu, Yan, Shen, Cheng, Huang, Yao and Song2022), and potential anticancer properties (Hang et al. Reference Hang, Wu, Wang, Sheng, Fang and Guo2022).
In grassland ecosystems, S. chamaejasme poses a significant challenge. Its robust root system, competitive strength, and high biomass allow it to outcompete other forage grasses for nutrients and space, inhibiting their growth. This negatively impacts grassland productivity and sustainable animal husbandry (Ninan and Inoue, Reference Ninan and Inoue2013). From 1970 to 2017, invasive plants caused direct global economic losses totaling $100.468 billion (Zhao et al. Reference Zhao, Gao, Wang, Fan, Tang, Sun and Wen2024). In China, (The Third National Land Survey of the State Council, Reference Guo, Cui, Jin, Yan, Ding and Qin2021) Annual Bulletin of Natural Resources of China (2024). where grasslands cover approximately 26.45 million ha (27.5% of the total land area), more than 90% of these grasslands have already experienced degradation due to factors like improper grazing and climate change (Sun et al., Reference Sun, Tang, Yang, Liao, Song and Wang2024). Specifically, the toxic invasive plant S. chamaejasme has spread across 1.4 million ha of natural grasslands in Qinghai Province, 546,700 ha in Gansu Province, 133,000 ha in Ar Horqin Banner of Inner Mongolia, and 32,300 ha in Zhaosu County, Ili Prefecture, Xinjiang. Taking Gansu Province as an example, more than 466,700 ha of grassland have been affected by this species, leading to a loss of 137,500 Mg of grazing forage and economic damages estimated at 15 to 20 million yuan (Wang et al. Reference Wang, Ma, Geng, Feng, Wu, Wang and Zhao2015). Its biological characteristics and potent toxicity act as a protective barrier, deterring herbivores from consuming it, thereby granting it a competitive advantage over neighboring plants (He et al. Reference He, Detheridge, Liu, Wang, Wei, Griffith, Scullion and Wei2019; Li et al. Reference Li, Xiang, Tang, Jiang, Duang and Chang2019). Its allelopathic effects further contribute to its dominance (Song et al. Reference Song, Li, Cheng, Song and Huang2023). The phenomenon of S. chamaejasme rapidly invading, expanding, and establishing dominance in grassland ecosystems due to its advantageous traits (Figure 1) has occurred in grasslands in different regions (He et al. Reference He, Detheridge, Liu, Wang, Wei, Griffith, Scullion and Wei2019; Javed et al. Reference Javed, Sun, Azeem, Ullan, Huang, Kama, Jabran and Du2019; ZY Wang et al., Reference Wang, Liu, Zhou and Li2022; You et al. Reference You, Ma, Guo, Kong, Shi, Wu and Zhao2018).

Figure 1. Stellera chamaejasme invades different grasslands in the Qilian Mountains. (A–C) Habitat: Kang le Grassland, Shan dan Military Horse Farm in Zhangye, City Xi shui Nature Reserve Station; (D–F) drone images; (G–I) individual plants.
The widespread distribution of S. chamaejasme, spanning from southern Russia to southwestern China and the western Himalayas, contributes to the prevalence of this phenomenon (CZ Zhang et al. Reference Zhang, Volis and Sun2010; YH Zhang et al. Reference Zhang, Yue and Sun2015). Its impact on grassland ecosystems is severe, not only reducing productivity but also posing economic losses due to acute poisoning in livestock that consume it (Bellard et al. Reference Bellard, Cassey and Blackburn2016; Murphy et al. Reference Murphy and Romanuk2014). Understanding the mechanisms behind this phenomenon is crucial for effective control measures and the restoration of degraded grasslands to support biodiversity conservation. This review aims to conduct a comprehensive description of the impacts of S. chamaejasme on plant communities and herbivore interactions, elucidating how this species successfully invades grasslands and becomes a dominant species, thereby providing a scientific foundation for the development of management strategies.
Impacts on Plant Communities
General Impact of Stellera chamaejasme
Stellera chamaejasme significantly impacts local plant communities by altering the soil’s physicochemical properties, microbial community structure, and enzymatic activity. These changes not only reshape the soil environment but also indirectly affect the growth and distribution of other plants. This ecological interaction is not only beneficial for the growth of S. chamaejasme, but also accelerates its expansion, which is one of the reasons why it can successfully invade grasslands (Figure 2).
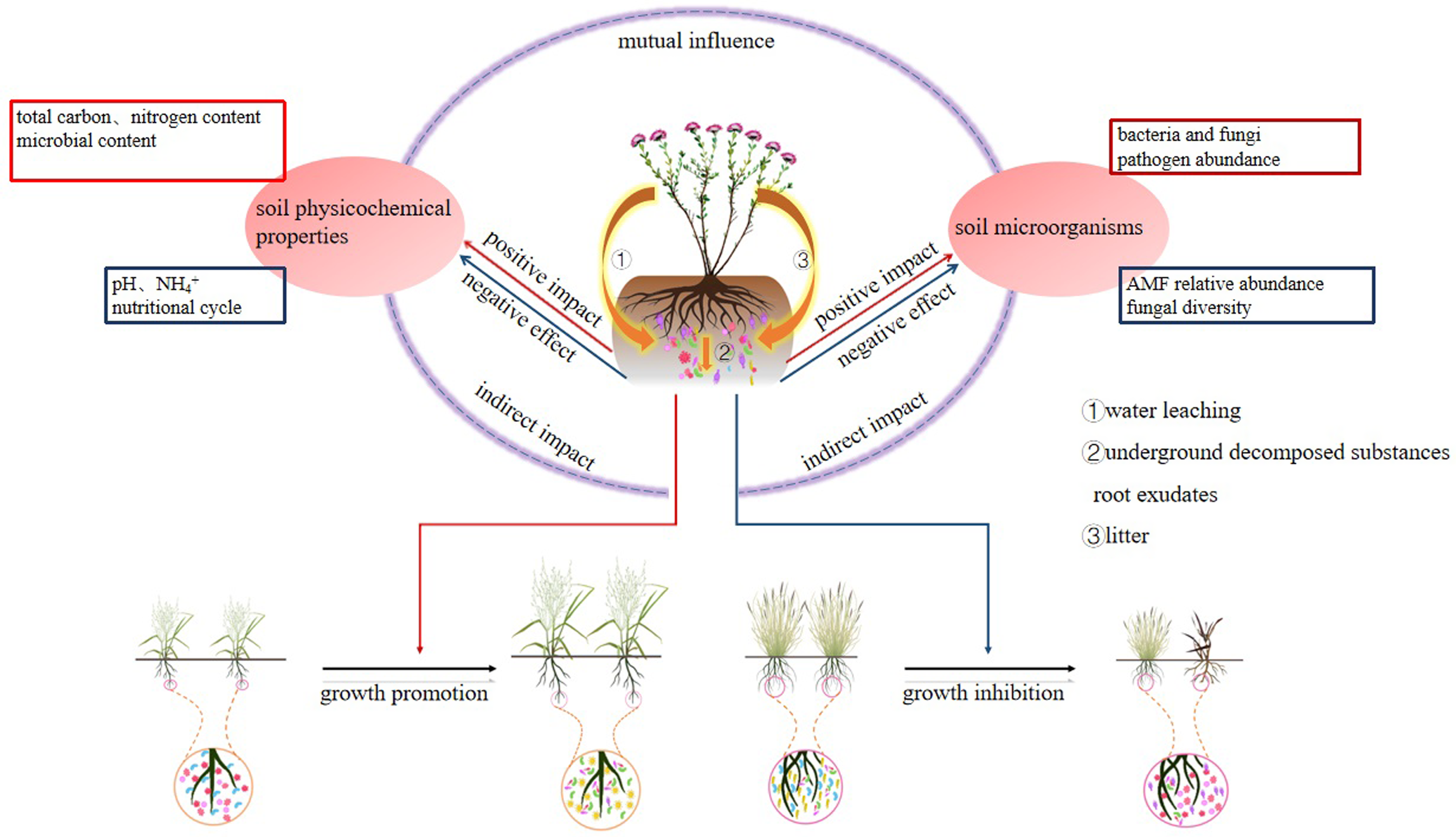
Figure 2. Mechanisms of Stellera chamaejasme impact on plants through soil factors. AMF, arbuscular mycorrhizal fungi.
Soil Physicochemical Properties
Studies have shown that extracts and plant residues of S. chamaejasme have significant impacts on the soil environment. They can reduce electrical conductivity, nitrogen, and available phosphorus while increasing soil pH and total carbon content (Zhu et al. Reference Zhu, Li, Xing, Chen, Huang and Gao2020). These alterations affect nutrient availability in the soil, subsequently influencing the growth of other plant species. Furthermore, as the S. chamaejasme community expands, soil microbes are affected by both nutrient depletion and secondary metabolites, leading to an increase in the content of organic matter, total nitrogen, and nitrate ions. This is conducive to the growth and spread of S. chamaejasme, while the levels of ammonium ions and soil pH are reduced (Y Zhang et al., Reference Zhang, Cui, Wang and Cao2021). These findings suggest that S. chamaejasme, through its released secondary metabolites, not only suppresses other plant growth but also improves soil conditions to suit its own growth requirements. Stellera chamaejasme litter, by regulating soil temperature and moisture, affects seed germination and establishment. In degraded grasslands, seeds that are exposed or shallowly buried may disrupt seedling establishment and population development (Xia et al. Reference Xia, Zhang, Li, Kong and Wang2021). Furthermore, similar to other invasive plants, S. chamaejasme’s expands and disrupts nutrient cycling in rhizosphere soil, leading to reduced soil nutrient content and affecting soil microbial activity (Lu-Irving et al. Reference Lu-Irving, Harenčár, Sounart, Welles, Swope and Baltrus2019).
Typically, a plant’s expansion exacerbates nutrient consumption from the external environment, resulting in soil impoverishment and decreased fertility. However, S. chamaejasme research reveals that it can alter soil nutrients through root exudates and litter, enhancing soil biological activity and accelerating nutrient transformation. This contributes to improved soil fertility, creating favorable conditions for S. chamaejasme growth and spread (An et al. Reference An, Han, Wu, Chen, Yuan, Liu and Wang2016; Dan et al. Reference Dan, Yi, Yang, Mei, Xie and Chu2021). Additionally, S. chamaejasme–inhabited soils exhibit higher organic nitrogen and microbial biomass, forming “fertility islands” that enhance its dispersal capacity (Crawford et al. Reference Crawford and Knight2017). Increased soil electrical conductivity may also promote S. chamaejasme invasion (Zhang et al. Reference Zhang, He, Xia, Lian, Yan, Ding, Zhang, Xu and Zhang2021).
Soil Microorganisms
Soil enzymes play a crucial role in soil ecosystems, participating in all carbon-, nitrogen-, and phosphorus-cycling processes (Hu et al. Reference Hu, Wang, Zhang, Shi, Jin and Chen2016). Therefore, studying enzyme activity in the rhizosphere soil of plants is essential for understanding the microbial environment. Research indicates that during its growth and expansion, S. chamaejasme significantly influences soil enzymes (An et al. Reference An, Han, Wu, Chen, Yuan, Liu and Wang2016). In the rhizosphere soil of S. chamaejasme during different growth seasons, enzyme activities related to sucrose, urease, and phosphatase are notably higher compared with vegetation-free soil. Interestingly, the activities of catalase and peroxidase vary during different growth stages of S. chamaejasme (An et al. Reference An, Han, Wu, Chen, Yuan, Liu and Wang2016). Additionally, Cheng et al. (Reference Cheng, Jin, Zhang, Xu, Yang, Liu, Xu, Min, Lu and Qin2022) found that secondary metabolites secreted by S. chamaejasme roots interact with soil enzymes. Notably, during the initial stages of S. chamaejasme coverage, the impact on soil enzyme activity is most pronounced. Rhizosphere bacteria play a vital role in nutrient cycling within soil ecosystems, significantly influencing plant health and soil fertility (Wu et al. Reference Wu, Hao, Zeng, Guo, Huang and Chen2015). As S. chamaejasme expands, it alters rhizosphere soil nutrient availability and bacterial community structure. The active participation of bacterial communities in carbon and nitrogen metabolism promotes plant growth under adverse conditions (Hu et al. Reference Hu, Sun, Wu, Liu, Jin and Tao2023). Research reveals that S. chamaejasme rhizosphere soil harbors a greater diversity of bacterial and fungal species compared with non-Stellera areas (Dan et al. Reference Dan, Yi, Yang, Mei, Xie and Chu2021). Notably, the fungal community structure in rhizosphere soil differs significantly from endophytic fungi (Cheng et al. Reference Cheng, Jin, Xu, Yang, Qin and Zhang2021; Ma et al. Reference Ma, Hou and Bowatte2019). Nakamura et al. (Reference Nakamura, Toju and Kitajima2023) found distinct differences in fungal and bacterial communities between invasive plant leaves, roots, soil, native, and exotic distributions. This suggests that invasive plants, including S. chamaejasme, may influence soil microbiota through root and leaf interactions, potentially favoring their own growth and expansion. Furthermore, Jin et al. (Reference Jin, Yang, Liu, Yan, Li and Li2018) discovered that S. chamaejasme rhizosphere and root-associated bacteria include Actinobacteria and Proteobacteria, which can produce hormones like gibberellins and auxins. These bacteria play essential roles in various processes, such as antibiotic production, bioactive compound synthesis, secondary metabolite production, phosphate solubilization, nitrogen fixation, and promoting plant growth (Gan et al. Reference Gan, Lin, Zhang and Bai2017; Sánchez-Gil et al. Reference Sánchez-Gil, Poppeliers, Vacheron, Zhang, Odijk, Keel and de Jonge2023). They also exhibit significant biocontrol effects against a range of plant pathogens (Anwar et al. Reference Anwar, Ali and Sajid2016). These interactions favor S. chamaejasme growth and expansion (Jin et al. Reference Jin, Guo, Yang, Xin, Liu and Qin2022b). Additionally, Bao et al. (Reference Bao, Song, Wang, Yin and Wang2020) reported that the biomass and diversity indices of S. chamaejasme patches were higher than those outside the patches. This aligns with Cui et al.’s (Reference Cui, Pan, Wang, Zheng and Gao2020) findings that high-density S. chamaejasme growth positively influenced microbial composition in rhizosphere soil, resulting in higher microbial diversity. Jin et al. (Reference Jin, Cheng, Liu, Yang, Dai, Huang, Yan, Min, Xu and Qin2024) explored S. chamaejasme and its companion grasses (Stipa purpurea Griseb. and Polygonum viviparum L.) and found that the S. chamaejasme rhizosphere tends to harbor dominant bacterial communities, enhancing microbial diversity and richness. Compared with associated grass (Leymus chinensis (Trin.) Tzvelev), the fungal community in the S. chamaejasme rhizosphere significantly decreased. Furthermore, the addition of S. chamaejasme litter positively impacted soil nutrient availability, although it had detrimental effects on arbuscular mycorrhizal fungi (AMF) in rhizosphere soil due to changes in nutrient balance and allelopathic compounds (Guo et al. Reference Guo, Zhang, Li, Liu, Li and Xing2022).
In their extensive investigation of microbial communities associated with three invasive weed species in North American grasslands, Gibbons et al. (Reference Gibbons, Lekberg, Mummey, Sangwan, Ramsey and Gilbert2017) discovered that invasive species can gain an advantage by altering soil community structure. Stellera chamaejasme has the ability to modify the soil nitrogen-fixing microbial community structure in grasslands, resulting in changes in the relative abundance of rhizobial species (Zhang et al. Reference Zhang, He, Xia, Lian, Yan, Ding, Zhang, Xu and Zhang2021). Furthermore, research indicates that in soils invaded by S. chamaejasme, nitrogen, phosphorus, organic matter, fungal alpha diversity, and AMF relative abundance are lower, while pathogenic fungi abundance is higher (He et al. Reference He, Detheridge, Liu, Wang, Wei, Griffith, Scullion and Wei2019). In plant communities, a high relative abundance of fungal pathogens can negatively impact plant growth. However, S. chamaejasme contains flavonoids such as neochamaejasmin B, chamechromone, and isochamaejasmin, which can protect the plant from these pathogens (ZQ Yan et al. Reference Yan, Zeng, Jin and Qin2015). Consequently, in soils with a high incidence of pathogens, S. chamaejasme may have a competitive advantage over coexisting species. This phenomenon is reminiscent of Canada thistle [Cirsium arvense (L.) Scop.], which influences local plant growth through soil microbiota pathways (such as pathogen accumulation), promoting its own expansion (Verbeek et al. Reference Verbeek and Kotanen2019). Dominant plant species can gain a competitive growth advantage by modifying the soil microbial composition through increasing pathogen abundance. This alteration specifically inhibits the normal growth of neighboring plants while allowing the invasive plants to remain unaffected due to their protective secondary metabolites that confer resistance against these pathogens. Tang et al. (Reference Tang, Gong, Xiao, Mao, Zhao, Song, Awais, Ji and Li2023) found that S. chamaejasme harbors a diverse endophytic fungal community, with most species exhibiting plant growth–promoting activity. This plays a crucial role in the rapid spread of S. chamaejasme in degraded grasslands. Additionally, a comparison of ammonia-oxidizing bacteria communities in S. chamaejasme and Dahurian wild rye (Elymus dahuricus Turcz. ex Griseb.) soils by Ma et al. (Reference Ma, Bowatte, Wang, Newton and Hou2020) revealed slower soil nitrification in S. chamaejasme soils compared with those under E. dahuricus. Reduced soil nitrification can lead to conservative nitrogen cycling, potentially conferring greater competitiveness to plants. This may partly explain why S. chamaejasme thrives as a dominant species in degraded grasslands.
Stellera chamaejasme significantly impacts soil ecosystems through secondary metabolites secreted by its roots and leaf litter. These compounds not only alter soil microbial community structure, influencing the growth of specific microorganisms, but also affect soil physicochemical properties such as pH, electrical conductivity, and nutrient content, consequently impacting nutrient availability and microbial survival in the soil.
Impacts through Resource Competition
Environmental Adaptability
As a perennial herbaceous plant, Stellera chamaejasme, possesses a well-developed root system that allows it to thrive even in adverse conditions. The roots can penetrate deep into the soil (with depths ranging from 10 cm for 1-yr-old plants to 60 to 100 cm for 4-yr-old plants), enabling efficient nutrient absorption (Li et al. Reference Li, Xiang, Tang, Jiang, Duang and Chang2019). Zhang et al. (Reference Zhang, Wu, Wang, Dai and Peng2024) highlighted that certain root system characteristics, such as a deep root architecture, play a crucial role in crops’ effective absorption of water and nutrients under arid conditions. The root system of S. chamaejasme aligns with this trait, suggesting that a developed root structure is beneficial for plants to adapt to adverse environments. Additionally, some invasive plants outperform native species in nutrient acquisition through their root systems (Chen et al. Reference Chen, Zhang, Liu, Han and Kong2022). Thus, S. chamaejasme’s robust root system not only supports its growth but also facilitates successful establishment and spread in new environments.
Stellera chamaejasme’s lanceolate leaves are covered by a thick waxy layer, enabling leaves to withstand prolonged drought. Unlike other plants that wither under extreme dry conditions, S. chamaejasme remains lush and vigorous (Lee et al. Reference Lee and Suh2015). The narrow shape of its leaves, combined with the waxy surface, significantly reduces water loss, enabling the plant’s survival in arid environments (J Guo et al. Reference Guo, Xu, Yu, Shen, Li, Cheng, Liu, Liu, Liu, Zhao and Song2016). Consequently, S. chamaejasme exhibits strong tolerance to adverse conditions, allowing it not only to survive but also to reproduce rapidly and spread. Research suggests that invasive species often display superior drought resistance compared with native plants, which contributes to their competitive advantage (Javed et al. Reference Javed, Sun, Azeem, Ullan, Huang, Kama, Jabran and Du2019; Sun et al. Reference Sun, Javed, Du, Azeem, Abbas, Lqbal, He, Xiang and Du2022).
As altitude increases, S. chamaejasme exhibits correlated changes in flower and leaf size and quantity. This reflects its adaptive strategies during long-term interactions with diverse environments, allowing it to thrive even in unstable growth conditions (Zhang et al. Reference Zhang, Zhao, Dong, Ma, Hou and Li2014). These traits align with the patterns observed in invasive species within plant communities. Invasive species often maximize light capture and photosynthetic efficiency through larger branches and higher specific leaf area. They also allocate more nitrogen to photosynthesis-related processes, enhancing their competitive advantage (Dyderski et al. Reference Dyderski and Jagodziński2019; Musso et al. Reference Musso, Fontenle, Pinto, Oliveira, Correia, Moutinho-Pereira, Soares and Loureiro2021). Stellera chamaejasme’s resilience after drought further contributes to its success in competitive environments (Zhang et al. Reference Zhang, Oduor and Liu2023).
In the context of ecological adaptation, S. chamaejasme exhibits remarkable adaptability in both root characteristics and leaf traits. Furthermore, it demonstrates resilience in adverse environments. These features not only underscore the plant’s high environmental adaptability but also reflect its endurance under challenging conditions. Consequently, even when soil nutrients are disrupted by external factors such as human activities or climate change, S. chamaejasme maintains normal growth, development, and physiological functions. Moreover, this adaptability ensures a competitive advantage for S. chamaejasme in resource-limited environments, facilitating its sustained growth and expansion on degraded grasslands. Ultimately, this phenomenon contributes to the occurrence and enhancement of “grassland poisoning” by the S. chamaejasme plant
High Reproductive Capacity
Stellera chamaejasme forms clumps with multiple stems. At the top of each stem, it produces crown-shaped inflorescences. This plant exhibits robust seed production, with mature individuals yielding around 200 seeds per plant (Zhao et al. Reference Zhao and Wang2011). This prolific seed production capability facilitates extensive reproduction and rapid spread of S. chamaejasme (Li et al. Reference Li, Xiang, Tang, Jiang, Duang and Chang2019). Notably, these seeds possess remarkable resilience, even when subjected to adverse conditions over several years. The protective features of S. chamaejasme seeds contribute to their survival. Their hard, thick seed coat shields them from external pressures such as physical damage, cold, and drought (Li et al. Reference Li, Xiang, Tang, Jiang, Duang and Chang2019; Wu et al. Reference Wu, Liu, He and Chen2014). Sharma et al. (Reference Sharma, Singh, Garai, Rahaman, Khatun, Ranjan, Mishra and Tiwari2022) found that invasive species often exhibit high expansion potential due to their ability to produce abundant, long-lasting viable seeds in the soil.
The species relies on seed dispersal for population expansion. Its dispersal pattern is characterized as “near-mother dispersal” (Li Reference Li1995), whereby seeds typically fall around the mother plant, forming seed clusters. Subsequently, external forces cause seed displacement within the soil surface, gradually dispersing or homogenizing them. Notably, S. chamaejasme lacks long-distance dispersal ability (Luo et al. Reference Luo, Zhang and Fang2021). All the plant parts of S. chamaejasme (root, stem, leaf, and flower) are toxic. This toxicity serves as an effective natural defense mechanism, deterring herbivores from consuming it (Zhao et al. Reference Zhao, Gao, Wang, He and Han2013). Interestingly, despite its overall toxicity, S. chamaejasme still attracts pollinators. Research indicates a mutualistic relationship between S. chamaejasme and flower thrips (Thysanoptera), in which pollination success is limited by pollinator availability rather than pollen availability (Zhang et al. Reference Zhang, He, Xia, Lian, Yan, Ding, Zhang, Xu and Zhang2021). Therefore, the pollen dispersal of S. chamaejasme relies not only on traditional external forces (wind, water, animal activity) but also benefits from unique biological traits that provide additional pathways for dissemination. As a toxic plant, S. chamaejasme faces a lower risk of herbivore consumption, which to some extent limits its reproductive opportunities (as animals typically avoid pollinating toxic plants). However, S. chamaejasme naturally circumvents this limitation. Its toxicity not only reduces the likelihood of herbivore consumption but also attracts specific pollinators, providing robust support for its reproduction and spread. The clumped growth structure of the stems allows S. chamaejasme to dominate spatially, reducing competition with other plants. Additionally, the crown-shaped inflorescences enhance pollination efficiency, significantly increasing the chances of successful pollination (Huang et al. Reference Huang, Jin, Li, Zhang, Yang and Wang2014). Despite its seed dispersal mode being near-mother distribution, this localized dispersal benefits the growth of S. chamaejasme seedlings. The soil conditions around the mother plant often support seed germination and seedling growth, promoting healthy seedling development (Peng et al. Reference Peng, Huang, Peng, OuYang and Xu2004). Furthermore, mature S. chamaejasme plants provide protection to seedlings during their growth (Wang et al. Reference Wang, Wang, Ren, Dou, Miao, Liu, Huang and Wang2022). The species’ ability to rapidly produce numerous offspring gives it a competitive advantage, allowing efficient acquisition of critical resources such as light, water, and nutrients. Moreover, its rapid reproductive and dispersal capacity enables it to spread to broader geographic areas within a short time. This not only facilitates rapid colonization of ecological niches and spaces in new locations but also helps establish stable populations.
High Genetic Diversity
Genetic analysis of the chloroplast genome of S. chamaejasme reveals unique adaptive evolutionary features compared with closely related species (Yun et al. Reference Yun, Park and Oh2019). Using amplified fragment length polymorphism markers, researchers have explored the genetic diversity and population structure of S. chamaejasme. The results indicate significant genetic differentiation among populations and relatively low gene flow, suggesting that this species is developing distinct genetic traits to adapt to its environment (Zhang et al. Reference Zhang, Yue and Sun2015). Rana et al. (Reference Rana, Rana, Landis, Kuang, Chen, Wang, Deng, Davis and Sun2024) investigated the effects of Quaternary climate change and glaciation on the genetic diversity of S. chamaejasme, emphasizing the role of high altitude and environmental heterogeneity in the development of cryptic species within it. Their research sheds new light on the plant’s high genetic diversity and the interplay between this diversity and environmental heterogeneity. This interplay allows the plant to adapt to varied environments and may explain its rapid spread on degraded grasslands. Similarly, another invasive plant, East Asian buttercup (Ranunculus japonicus Thunb.), also exhibits high genetic diversity, enabling it to maintain stable populations even under adverse conditions such as increasing altitude and environmental degradation (Yu et al. Reference Yu, Chen, Chen, Wei, Song, Liu, Liu, Li and Liu2023). Additionally, real-time quantitative polymerase chain reaction analysis of S. chamaejasme demonstrates stable gene expression under various abiotic stress and hormone treatments, further explaining its adaptive capacity to environmental changes (Liu et al. Reference Liu, Guan, Song, Fu, Han, Lei, Ren, Guo, He and Wei2018).
Studies have also revealed floral color polymorphism in S. chamaejasme. High genetic diversity (h(T) = 0.834) and population differentiation (N(ST) = 0.997 and G(ST) = 0.982) indicate restricted gene flow among populations, reflecting significant geographic or environmental isolation. This underscores S. chamaejasme’s remarkable adaptability to different environments (Zhang et al. Reference Zhang, Volis and Sun2010). Recent research further reveals four distinct genetic lineages of S. chamaejasme in the Qinghai-Tibet Plateau and northern China, corresponding to different environmental distributions. Highly divergent adaptive evolution genes are widely distributed both within and outside genomic islands, closely associated with local adaptation (Hu et al. Reference Hu, Yang, Li, Zheng, Zhang and Liu2022). Moreover, studies by Wan et al. (Reference Wan, Fu, Dainese, Hu, Kiær, Isbell and Scherber2022) demonstrate that increasing plant genetic diversity decreased the performance of plant antagonists, while increasing the performance of plants and natural enemies of herbivores. Similar findings in the study of seed production in Austrian dragonhead (Dracocephalum austriacum L.) by Dostálek et al. (Reference Dostálek, Münzbergová and Plačková2010) highlight the advantages of high genetic diversity.
Impacts through Allelopathy
Allelochemicals of Stellera chamaejasme
Guo et al. (Reference Guo, Cui, Jin, Yan, Ding and Qin2015) identified seven allelochemicals in the roots of S. chamaejasme (Figure 3), namely umbelliferone, chamaechromone, daphnoretin, mesoneochamaejasmin A, neochamaejasmin B, dihydrodaphnodorin B, and 7-methoxyneochamaejasmine A. Among these, five allelochemicals are found in higher concentrations in the roots and rhizosphere soil (Jin et al. Reference Jin, Cheng, Cheng, Yang, Guan and Qin2022a). Research indicates that umbelliferone may be the primary allelochemical in S. chamaejasme, and its degradation may play a crucial role in competition with other plants (Guo et al. Reference Guo, Cui, Jin, Yan, Ding and Qin2015).
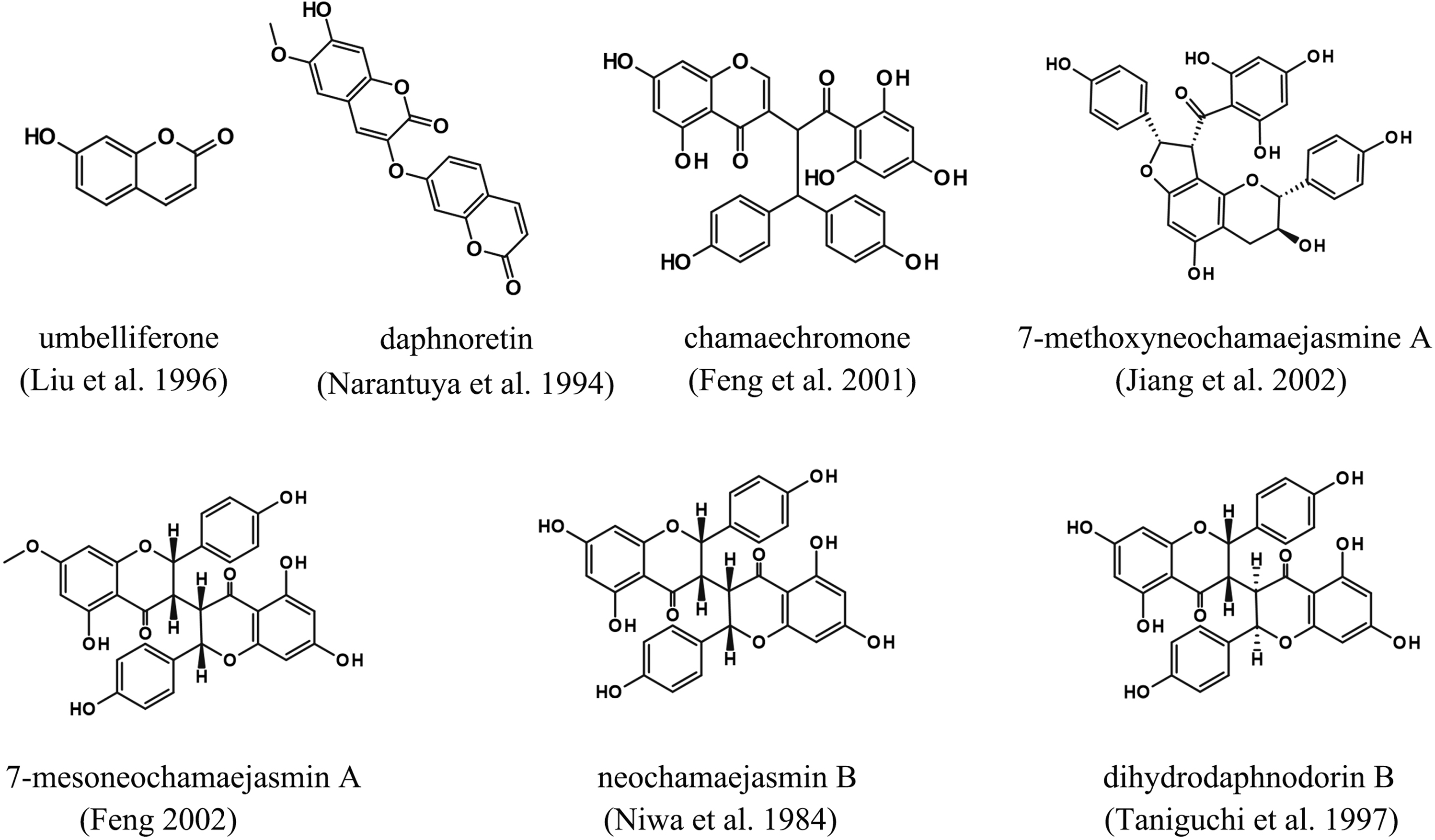
Figure 3. Main allelochemicals from Stellera chamaejasme. Sources: Feng Reference Feng2002; Feng et al. Reference Feng, Pei and Han2001; Jiang et al. Reference Jiang, Tanaka, Sakamoto, Kouno, Duan and Zhou2002; Liu et al. Reference Liu, Zhao, Hou, Yang, Zhao and Liu1996; Narantuya et al. Reference Narantuya, Batsurén, Rashkes and Mil’grom1994; Niwa et al. Reference Niwa, Tatematsu, Liu and Hirata1984; Taniguchi et al. Reference Taniguchi, Fujiwara and Baba1997.
Allelopathy of Stellera chamaejasme
Allelopathy is one of the important reasons why S. chamaejasme can spread on a large scale on grasslands. This term refers to the ability of plants to inhibit the growth of surrounding plants by releasing specific chemicals (Macías et al. Reference Macías, Mejías and Molinillo2019; K Wang et al. Reference Wang, Wang, Ren, Dou, Miao, Liu, Huang and Wang2022) into the growth medium. The pathways through which S. chamaejasme releases these allelopathic substances mainly include: (1) rainwater leaching (Fernandez et al. Reference Fernandez, Monnier, Santonja, Gallet, Weston, Prévosto, Saunier, Baldy and Bousquet-Mélou2016), (2) the decomposition of plant residues (Kong et al. Reference Kong, Xuan, Khanh, Tran and Trung2019), and (3) root exudates (Weston and Mathesius, Reference Weston and Mathesius2013: HR Guo et al. Reference Guo, Zeng, Yan, Jin, Li, Guan and Qin2016) (Figure 4). At present, research on the allelopathic effects of S. chamaejasme mainly focuses on simulating these release pathways to further explore the specific mechanisms of its allelopathic effects.
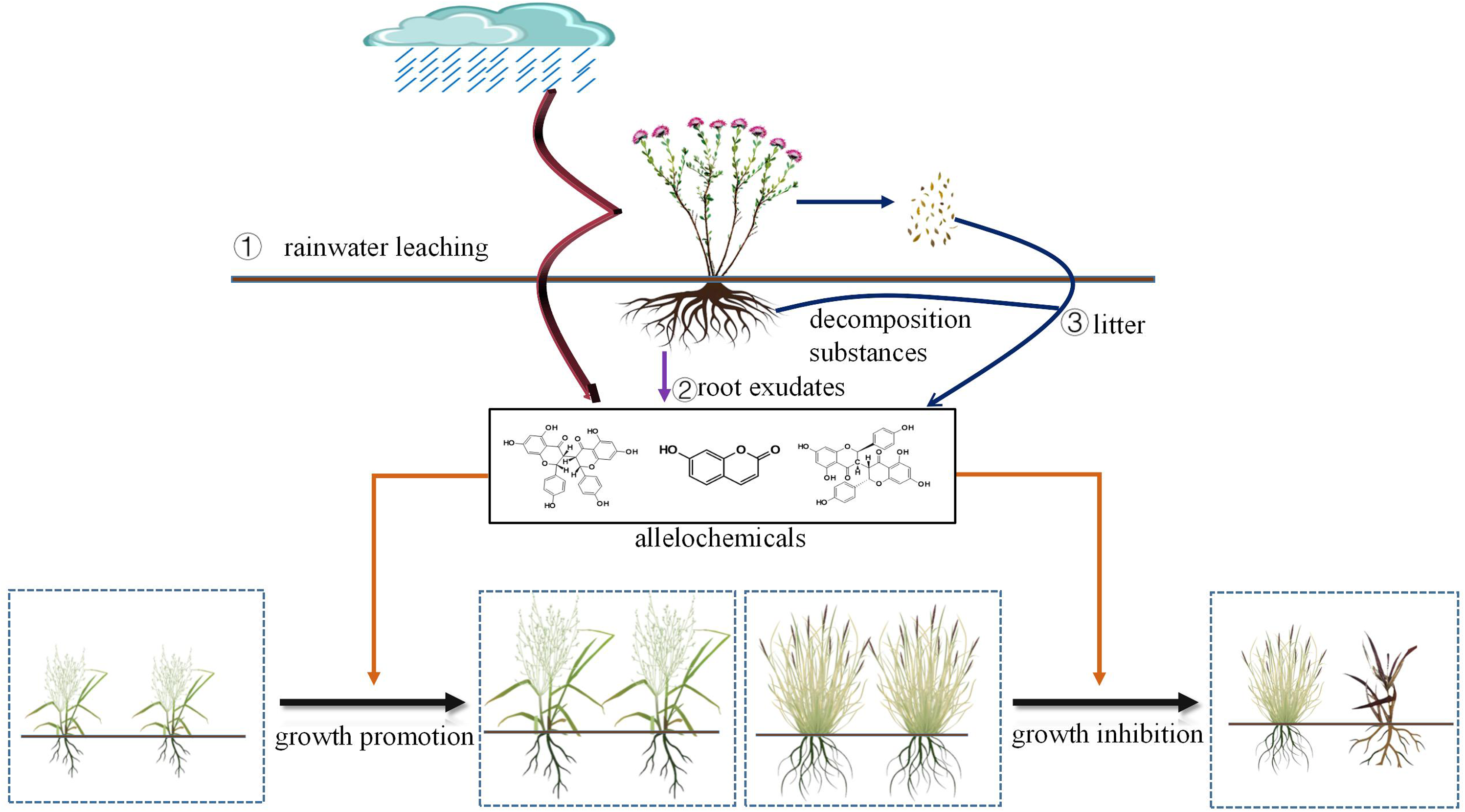
Figure 4. Release pathways and allelopathy mechanisms of allelochemicals from Stellera chamaejasme.
Stellera chamaejasme exhibits allelopathy through its roots, stems, leaves, and pollen. Several studies have simulated the release pathways of allelochemicals from S. chamaejasme and other plants, revealing that the allelochemical impact generally increases with the quantity of its root, stem, and leaf material. For instance, Wang et al. (Reference Wang, Zhou and Huang2009) found that S. chamaejasme’s allelopathy on sweetclover (Melilotus suaveolens Ledeb.) and perennial ryegrass (Lolium perenne L). intensified with increasing root, stem, and leaf biomass. Similarly, Wei et al. (Reference Wei, Wang, Wu, Cheng and Wang2020) observed enhanced allelopathy in lettuce (Lactuca sativa L.) as the concentration of Canada goldenrod (Solidago canadensis L.) leaf extracts increased. Notably, S. chamaejasme’s allelopathic impact on certain receptor plants exhibits a “low promotion, high inhibition” phenomenon (YJ Liu et al. Reference Liu, Meng, Dang, Song and Zhuo2019; GZ Liu et al. Reference Liu, Liu, Guo, Su, Lan and Liu2022). Studies on alfalfa (Medicago sativa L.) and Daurian wild rye (Elymus dahuricus) revealed that the allelopathy of Euphorbia jolkinii displayed this pattern (Li et al. Reference Li, Chu, Li, Ma, Niu and Dan2022).
Cheng et al. (Reference Cheng, Zhong, Xu, Du and Song2017) investigated high-altitude alpine meadow plants on the Qinghai-Tibet Plateau and found that S. chamaejasme root extracts generally exhibited stronger allelopathy than stem and leaf extracts. This aligns with the results from Wang et al. (Reference Wang, Wang, Ren, Dou, Miao, Liu, Huang and Wang2022) in alpine meadows. However, other studies suggest that S. chamaejasme stem and leaf allelopathy may surpass that of the roots. For instance, Liu et al. (Reference Liu, Liu, Guo, Su, Lan and Liu2022) discovered that high-concentration S. chamaejasme stem leaf infusions had a stronger inhibitory effect on German garlic (Allium senescens Thunb.) seed germination and seedling growth compared with root infusions. These differences may be related to varying sensitivities of receptor plants to allelochemicals, resulting in inconsistent responses to the same allelopathic substance. Additionally, Sun et al. (Reference Sun, Luo and Wu2010) found that S. chamaejasme pollen extracts significantly inhibited seed germination and fruit set. Similarly, the pollen of invasive plants like jewelweed (Impatiens glandulifera Royle.) can also exhibit allelopathic effects, inhibiting neighboring plant seed germination and growth (Beans and Roach Reference Beans and Roach2015). Furthermore, Cheng et al. (Reference Cheng, Zhong, Xu, Du and Song2017) observed that S. chamaejasme’s allelopathic effects were positively correlated with plant age, suggesting that allelochemical content generally increases with plant age and decreases with organ senescence.
Elymus dahuricus and crested wheatgrass [Agropyron cristatum (L.) Gaertn.] exhibit some tolerance to S. chamaejasme allelopathy, making them suitable grass species for restoring grassland vegetation invaded by S. chamaejasme (Liu et al. Reference Liu, Meng, Dang, Song and Zhuo2019). This finding highlights differences in allelopathy across different S. chamaejasme plant parts and varying tolerance levels among different plant species. Uddin et al. (Reference Uddin, Asaeda, Shampa and Robinson2020) also observed similar phenomena in interactions between native and invasive plants, suggesting coevolution of tolerance. This implies that the relationship between S. chamaejasme and neighboring plants may involve a coexistence pattern rather than unidirectional competition. Furthermore, the intensity of S. chamaejasme’s allelopathy depends not only on the source organ (roots, stems, or leaves) but also on the specific allelochemical type. This also explains why the S. chamaejasme can gradually transition from being a “companion species” to being a “dominant species” in grasslands without completely replacing other plants.
Impact of Herbivores on Stellera chamaejasme
Research indicates that herbivore diversity in grassland ecosystems, including grazing livestock, wildlife, and insects (Gao et al. Reference Gao, Wang, Cheng, Kang, Ma, Han and Li2022; Yue et al. Reference Yue, Gao, Cheng, Xu, Han, Ding and Chai2021), has multifaceted effects. These herbivores play a role in both promoting the spread of invasive plants and driving grassland degradation, ultimately leading to S. chamaejasme becoming a dominant species (Figure 5). Huang et al. (Reference Huang, Carrillo, Ding and Siemann2012) found that invasive plants, compared with native species, suffer less damage from herbivores. This difference may be attributed to characteristics of their leaf composition, such as lower concentrations of cellulose, hemicellulose, lignin, and carbon, as well as lower leaf density and carbon-to-nitrogen ratio, all while maintaining higher nutrient content (Beans and Roach Reference Beans and Roach2015). Notably, herbivores do not always choose based on preference when faced with S. chamaejasme and other plants. Stellera chamaejasme is toxic throughout its entire plant, and grazing livestock may be poisoned by inhaling its pollen or experiencing allergic dermatitis upon contact. The toxic compounds, which include neochamaejasmin B, umbelliferone, and chamaechromone, generally encourage livestock to avoid consuming fresh S. chamaejasme (Gillmeister et al. Reference Gillmeister, Ballert, Raschke, Geistlinger, Kabrodt, Baltruschat, Deising and Schellenberg2019; Yan et al. Reference Yan, Guo, Yang, Liu, Jin, Xu, Cui and Qin2014; You et al. Reference You, Ma, Guo, Kong, Shi, Wu and Zhao2018). However, under extreme hunger conditions in early spring, livestock may accidentally ingest S. chamaejasme seedlings and become poisoned. The selective pressure exerted by herbivores on S. chamaejasme via consumption leads to the depletion of valuable forage grasses, disruption of the reproductive cycle of perennial grasses, and reduced valuable forage grasses seedbank density (Gao et al. Reference Gao, Wang, Cheng, Kang, Ma, Han and Li2022; Li et al. Reference Li, Rahman, Zhu and Zhang2018). Meanwhile, the seedbanks of toxic weeds such as S. chamaejasme now dominate, making the restoration of degraded grasslands significantly more challenging (Zhao et al. Reference Zhao, Gao, Wang, Sheng and Shi2010). Surprisingly, despite its toxicity, S. chamaejasme’s growth and spread are indirectly benefited by reducing herbivore consumption, resulting in increased organic carbon content in the soil microaggregates around its roots (Zhou et al. Reference Zhou, Dong, Wang, Liu, Feng, Yang, Yu, Zhang, Cao and Liu2023). Cheng et al. (Reference Cheng, Sun, Du, Wu, Zheng, Zhang, Liu and Wu2014) discovered that S. chamaejasme not only protects itself from herbivore interference but also serves as a biological refuge for neighboring plants, safeguarding plant diversity from excessive grazing by livestock.
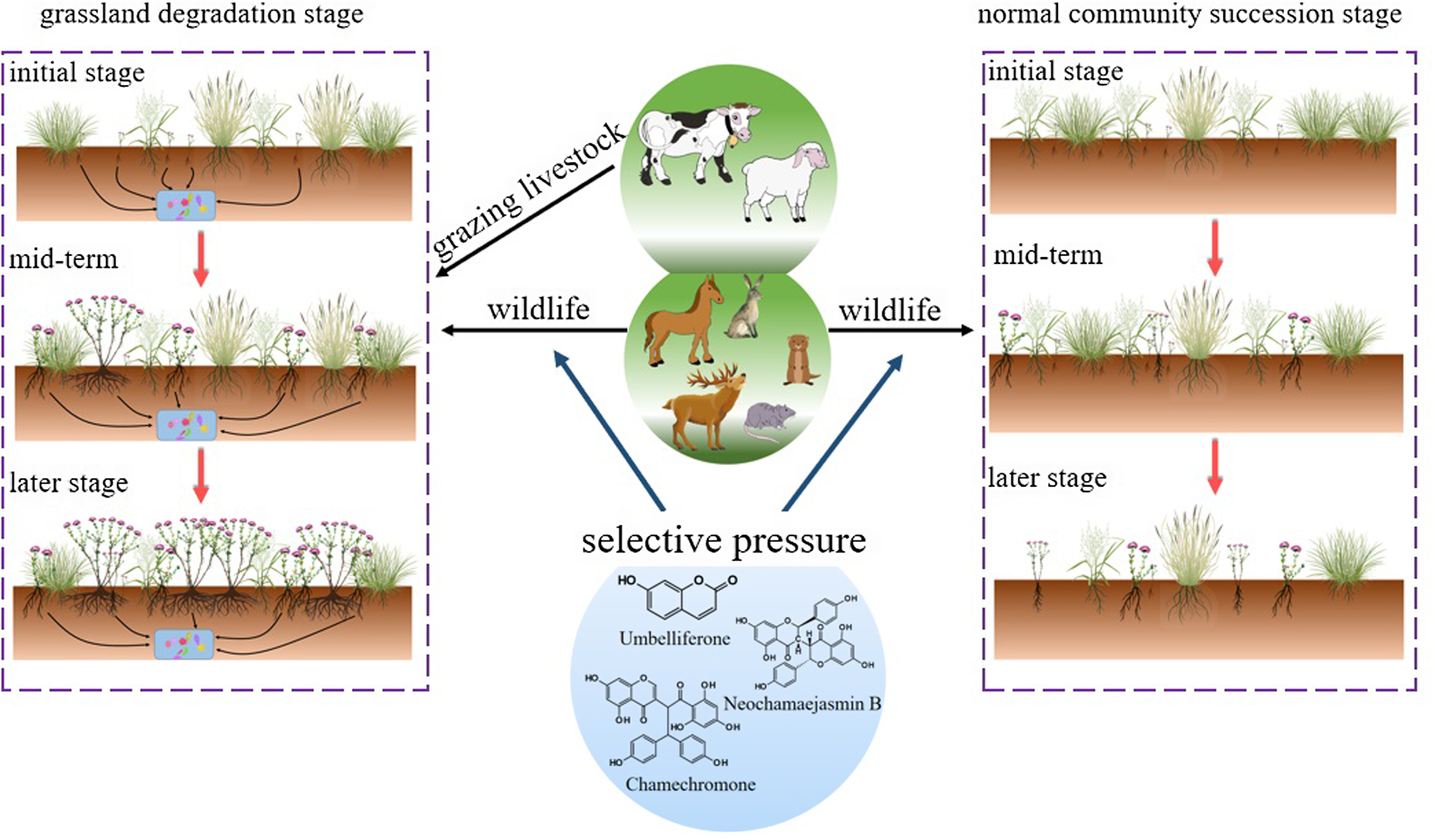
Figure 5. Dynamic changes in Stellera chamaejasme community succession under herbivore selective pressure.
The impact of herbivores on S. chamaejasme population distribution and expansion is complex. During heavy grazing or overgrazing, S. chamaejasme tends to exhibit aggregated distribution patterns, likely due to mutual protection within these clusters, allowing the population to withstand external pressures and stabilize (Zhao et al. Reference Zhao, Gao, Wang, Sheng and Shi2010). However, as grazing intensity increases, grassland degradation intensifies, and S. chamaejasme transitions from a companion species to a dominant one. Consequently, its distribution pattern may shift from aggregated to random or uniform, reflecting adaptive responses and ecological niche expansion (Gao et al. Reference Gao, Zhao, Shi, Sheng, Ren and He2011; Zhao and Ren Reference Zhao and Ren2011). Early-stage S. chamaejasme expansion typically involves aggregated distribution within patches centered around mature individuals, with random or uniform distribution within patches. As the number of mature individuals increases, intraspecific competition becomes dominant (Ren and Zhao Reference Ren and Zhao2013). Grazing activities may accelerate this transition, promoting patch enlargement, fusion, and an overall increase in patch numbers, driving S. chamaejasme population spread (Ren et al. Reference Ren, Nie, Jia, Gao, Li, Zhao, Li and Li2019; Ren and Zhao Reference Ren and Zhao2013). These findings highlight the intricate interplay between herbivore activity and S. chamaejasme population dynamics, which is crucial for understanding S. chamaejasme’s ecological spread in grassland ecosystems.
Conclusion
This article outlines the factors contributing to the successful invasion and subsequent dominance of S. chamaejasme in grassland ecosystems. By releasing secondary metabolites, S. chamaejasme modifies the soil environment, thereby reinforcing its dominant status. With a robust vitality and an exceptional reproductive capacity, S. chamaejasme has secured a competitive edge within these ecosystems. Notably, S. chamaejasme exhibits significant allelopathy, secreting allelochemicals through three main pathways that affect the growth and development of surrounding plants. Its strong allelopathic effects on a broad spectrum of plants endow S. chamaejasme with formidable invasive and expansion capabilities in novel environments. By inhibiting the growth of neighboring flora, it diminishes the diversity of species within the ecosystem, which in turn aids in its own propagation and strengthens its competitive position. The toxic properties of S. chamaejasme also reduce herbivore consumption, easing natural control pressures and enabling widespread growth and swift expansion, which in turn accelerate its invasion. As a result, the characteristics and behaviors of S. chamaejasme present considerable challenges to the health and stability of grassland ecosystems. To address this challenge, future research should focus on several key areas: (1) The sequential relationship between invasion and grassland degradation: undertake a systematic investigation to determine whether grassland degradation is a prerequisite for S. chamaejasme invasion or whether the invasion itself triggers degradation. (2) Ecological adaptation and allelopathic mechanisms: carry out an in-depth exploration of S. chamaejasme’s ecological adaptation mechanisms, including its responses to various environmental stressors and the specific pathways and mechanisms of allelopathic substance release. (3) Long-term effects on soil environment: investigate the long-term effects of S. chamaejasme on soil physicochemical properties and microbial community structure. Consider changes in soil nutrient cycling, microbial diversity, and enzyme activity. (4) Herbivore–plant interactions: systematically study the impact of herbivores on S. chamaejasme population dynamics and potential effects of S. chamaejasme on herbivore behavior. This includes herbivore preferences, seasonal feeding behavior, and their influence on S. chamaejasme growth and reproduction. (5) Integrated management strategies: develop and evaluate comprehensive management strategies for S. chamaejasme, including biological control, chemical treatments, and ecological restoration methods. These strategies should consider cost-effectiveness, sustainability, and potential ecosystem impacts. (6) Ecological restoration and biodiversity conservation: research techniques for restoring grassland ecosystems invaded by S. chamaejasme and enhancing biodiversity and ecosystem resilience. This involves selecting appropriate native plant species for vegetation reconstruction and assessing the long-term effects of different restoration measures. Through in-depth research on these critical issues, we can more effectively control the spread of S. chamaejasme, promote the restoration of degraded grasslands, and protect and maintain the health and stability of the grassland ecosystem.
Data availability
Data will be made available on request.
Funding statement
No funding was received for this work.
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.






 Open access
Open access