

1 Introduction
Many organisms experience periodic environmental variations influenced by seasonal changes and cyclical human activities. These fluctuations often affect the availability of resources (such as food, water, nutrients, and energy) as well as territories and habitats. For instance, seasonal succession (such as the succession of phytoplankton species [Reference Hsu and Zhao9]) and cyclical human behaviours (such as periodic open-closed fishery management policies [Reference Feng, Liu, Ruan and Yu6, Reference Liu, Feng, Ruan and Yu11–Reference Liu, Yu and Li13, Reference Xiao23] and the periodic release of Wolbachia-infected mosquitoes [Reference Yu and Li24, Reference Zheng, Zhou and Yu25, Reference Zheng, Yang, Elaydi and Yu26]) can cause the living environments of the populations to undergo regular changes. These periodic variations have profound effects on population growth dynamics and the success of species invasions.
The growth of organisms requires specific environmental conditions and hence not all the times are suitable for the growth of certain species. As a result, growth seasons alternate with non-growth seasons, during which biomass increases continuously and decreases gradually, respectively. For example, climate patterns repeat annually, birds migrate according to seasonal successions, and the opening and closing of schools are almost always cyclical. The incidence and contact rates of many infectious diseases are often influenced by seasonal changes. Autumn and winter are typically the high-incidence seasons for influenza. Childhood diseases such as measles, rubella, and whooping cough are highly sensitive to the seasonal pressures caused by the alternation between school terms and holidays. Seasonal factors significantly affect the survival of pathogens in the environment, host behaviours, and the abundance of vectors and non-human hosts. Existing extensive studies [Reference Augeraud-Véron and Sari1, Reference Barrientos, Rodríguez and Ruiz-Herrera2, Reference Duarte, Januário and Martins4, Reference Tang, Xiao, Zhang and Zhu20] suggest that seasonality plays a crucial role in the outbreak and evolution of epidemics and the transmission of diseases.
Pan et al. [Reference Pan, Shu, Wang, Wang and Yu15] analysed the following switching dynamical system,
 \begin{equation} \begin{cases} \dfrac {{\rm d} x}{{\rm d} t}=xf(x),\,&t\in (nT,nT+\overline {T}], \\[5pt] \dfrac {{\rm d} x}{{\rm d} t}=xg(x),\,&t\in (nT+\overline {T},(n+1)T]. \end{cases} \end{equation}
\begin{equation} \begin{cases} \dfrac {{\rm d} x}{{\rm d} t}=xf(x),\,&t\in (nT,nT+\overline {T}], \\[5pt] \dfrac {{\rm d} x}{{\rm d} t}=xg(x),\,&t\in (nT+\overline {T},(n+1)T]. \end{cases} \end{equation}
It is worth noting that the functions
 $f$
and
$f$
and
 $g$
in (1.1) are autonomous functions, namely, they are independent of the time
$g$
in (1.1) are autonomous functions, namely, they are independent of the time
 $t$
. However, in practical applications to biological systems, it has been observed that
$t$
. However, in practical applications to biological systems, it has been observed that
 $f$
and
$f$
and
 $g$
actually depend on
$g$
actually depend on
 $t$
, with specific details to be provided in the examples in Section 3. As a result, in this paper, we consider the following switching dynamical system,
$t$
, with specific details to be provided in the examples in Section 3. As a result, in this paper, we consider the following switching dynamical system,
 \begin{equation} \begin{cases} \dfrac{{\rm d} x}{{\rm d} t}=xf(t,x,\alpha ),\,&t\in (nT,nT+\overline {T}], \\[5pt] \dfrac{{\rm d} x}{{\rm d} t}=xg(t,x,\alpha ),\,&t\in (nT+\overline {T},(n+1)T], \end{cases} \end{equation}
\begin{equation} \begin{cases} \dfrac{{\rm d} x}{{\rm d} t}=xf(t,x,\alpha ),\,&t\in (nT,nT+\overline {T}], \\[5pt] \dfrac{{\rm d} x}{{\rm d} t}=xg(t,x,\alpha ),\,&t\in (nT+\overline {T},(n+1)T], \end{cases} \end{equation}
with
 $x(0)\in \mathbb{R}_{+}=[0,+\infty )$
(Here and in the sequel, without mentioning,
$x(0)\in \mathbb{R}_{+}=[0,+\infty )$
(Here and in the sequel, without mentioning,
 $n=0$
,
$n=0$
,
 $1$
,
$1$
,
 $2$
,
$2$
,
 $\ldots$
).
$\ldots$
).
 $T$
and
$T$
and
 $\overline {T}$
are two important time points satisfying
$\overline {T}$
are two important time points satisfying
 $T\gt \overline {T}$
. The parameter
$T\gt \overline {T}$
. The parameter
 $\alpha$
is of particular interest in this study and hence it is highlighted here.
$\alpha$
is of particular interest in this study and hence it is highlighted here.
We assume that both
 $f(t,x,\alpha )$
and
$f(t,x,\alpha )$
and
 $g(t,x,\alpha )$
from
$g(t,x,\alpha )$
from
 $\mathbb{R}_{+}\times \mathbb{R}_{+}\times \mathbb{R}_{+}$
to
$\mathbb{R}_{+}\times \mathbb{R}_{+}\times \mathbb{R}_{+}$
to
 $\mathbb{R}$
are continuous,
$\mathbb{R}$
are continuous,
 $T$
-periodic in
$T$
-periodic in
 $t$
, and continuously differentiable in both
$t$
, and continuously differentiable in both
 $x$
and
$x$
and
 $\alpha$
. Then it is easy to show the existence and uniqueness of solutions to system (1.2). In fact, taking
$\alpha$
. Then it is easy to show the existence and uniqueness of solutions to system (1.2). In fact, taking
 $n=0$
, by a standard application of the existence theory of ordinary differential equations, one can obtain the existence and uniqueness of solutions on
$n=0$
, by a standard application of the existence theory of ordinary differential equations, one can obtain the existence and uniqueness of solutions on
 $(0,\overline {T}]$
. Then, similarly, we can prove the existence and uniqueness of solutions in
$(0,\overline {T}]$
. Then, similarly, we can prove the existence and uniqueness of solutions in
 $(\overline {T},T]$
with the initial data
$(\overline {T},T]$
with the initial data
 $x(\overline {T})$
. Continuing this way, we have the existence and uniqueness of solutions to system (1.2) on
$x(\overline {T})$
. Continuing this way, we have the existence and uniqueness of solutions to system (1.2) on
 $\mathbb{R}_+$
.
$\mathbb{R}_+$
.
The rest of the paper is organised as follows. In Section 2, we provide the detailed analysis on the dynamics of (1.2) under certain reasonable assumptions. Then the obtained theoretical results are illustrated with two applications in Section 3. The paper concludes with a brief summary.
2 The dynamics of system (1.2)
In this section, we develop the existence and stability of positive periodic solutions to system (1.2) under certain assumptions.
We first state some assumptions on
 $f$
and
$f$
and
 $g$
.
$g$
.
-
(H1)
 $f(t,0,\alpha )\geq f(t,x,\alpha )$
and
$g(t,0,\alpha )\geq g(t,x,\alpha )$
for
$x\gt 0$
.
$f(t,0,\alpha )\geq f(t,x,\alpha )$
and
$g(t,0,\alpha )\geq g(t,x,\alpha )$
for
$x\gt 0$
. -
(H2) There exist
$M_{1}$
and
$M_{2}$
such that
$f(t,x,\alpha )\leq 0$
for all
$x\geq M_{1}\gt 0$
and
$g(t,x,\alpha )\le 0$
for
$x\geq M_{2}\gt 0$
. -
(H3)
$ \begin{cases} f_{x}(t,x,\alpha )\lt 0 \quad & \mbox{for $t\in (nT,nT+\overline {T}]$ and $x\gt 0$,} \\ g_{x}(t,x,\alpha )\lt 0 &\mbox{for $t\in (nT+\overline {T},(n+1)T]$ and $x\gt 0$.} \end{cases}$










Denote
 $\digamma (T,\overline {T},\alpha ) =\int ^{\overline {T}}_{0}f(t,0,\alpha )\,{\rm d}t+\int ^{T}_{\overline {T}} g(t,0,\alpha )\,{\rm d}t$
. We also have the following assumption on
$\digamma (T,\overline {T},\alpha ) =\int ^{\overline {T}}_{0}f(t,0,\alpha )\,{\rm d}t+\int ^{T}_{\overline {T}} g(t,0,\alpha )\,{\rm d}t$
. We also have the following assumption on
 $\digamma$
.
$\digamma$
.
-
(H) The function
$\digamma (T,\overline {T},\alpha )$
satisfies at least one of the following three assumptions about parameters
$T\gt 0$
,
$\overline {T}\gt 0,$
and
$\alpha \gt 0$
.-
(a) For any fixed
$T\gt 0$
and
$\alpha \gt 0$
,
$\digamma (T,\overline {T},\alpha )$
is strictly monotonic in
$\overline {T}$
and
$\digamma (T,\overline {T},\alpha )=0$
has a unique non-negative root with respect to
$\overline {T}$
. -
(b) For any fixed
$\overline {T}\gt 0$
and
$\alpha \gt 0$
,
$\digamma (T,\overline {T},\alpha )$
is strictly monotonic in
$T$
and
$\digamma (T,\overline {T},\alpha )=0$
has a unique non-negative root with respect to
$T$
. -
(c) For fixed
$T\gt 0$
and
$\overline {T}\gt 0$
,
$\digamma (T,\overline {T},\alpha )$
is strictly monotonic in
$\alpha$
and
$\digamma (T,\overline {T},\alpha )=0$
has a unique non-negative root with respect to
$\alpha$
.
-






















In the following, we assume that the monotonicity in (H) is strictly increasing. For results in the case of monotonically decreasing, just reverse the directions of inequalities in conditions. The non-negative root in (H) will serve as the threshold value. In fact, the linearisation of (1.2) at
 $0$
is
$0$
is
 \begin{equation} \begin{cases} \dfrac{{\rm d} y}{{\rm d} t}=yf(t,0,\alpha ),\quad &t\in (nT,nT+\overline {T}], \\[8pt] \dfrac{{\rm d} y}{{\rm d} t}=yg(t,0,\alpha ),&t\in (nT+\overline {T},(n+1)T]. \end{cases} \end{equation}
\begin{equation} \begin{cases} \dfrac{{\rm d} y}{{\rm d} t}=yf(t,0,\alpha ),\quad &t\in (nT,nT+\overline {T}], \\[8pt] \dfrac{{\rm d} y}{{\rm d} t}=yg(t,0,\alpha ),&t\in (nT+\overline {T},(n+1)T]. \end{cases} \end{equation}
By the Floquet theory, any fundamental solution of equation (2.1) has the form
 $y(t)=e^{-\lambda t}\psi (t)$
, where
$y(t)=e^{-\lambda t}\psi (t)$
, where
 $\lambda \in \mathbb{C}$
is a characteristic exponent and
$\lambda \in \mathbb{C}$
is a characteristic exponent and
 $\psi (t+T)=\psi (t)$
for all
$\psi (t+T)=\psi (t)$
for all
 $t\in \mathbb{R}$
. Thus, we have
$t\in \mathbb{R}$
. Thus, we have
 \begin{equation} \begin{cases} \dfrac{{\rm d} \psi }{{\rm d} t}=f(t,0,\alpha )\psi +\lambda \psi , \quad & t\in (0,\overline {T}], \\[8pt] \dfrac{{\rm d} \psi }{{\rm d} t}=g(t,0,\alpha )\psi +\lambda \psi ,& t\in (\overline {T},T], \\[8pt] \psi (0)=\psi (T) \end{cases} \end{equation}
\begin{equation} \begin{cases} \dfrac{{\rm d} \psi }{{\rm d} t}=f(t,0,\alpha )\psi +\lambda \psi , \quad & t\in (0,\overline {T}], \\[8pt] \dfrac{{\rm d} \psi }{{\rm d} t}=g(t,0,\alpha )\psi +\lambda \psi ,& t\in (\overline {T},T], \\[8pt] \psi (0)=\psi (T) \end{cases} \end{equation}
or equivalently,
 \begin{equation} \begin{cases} \dfrac {1}{\psi(t)} \dfrac{d\psi (t)}{dt}=f(t,0,\alpha )+\lambda , \quad & \mbox{ $t\in (0,\overline {T}]$,} \\[8pt] \dfrac {1}{\psi(t)}\dfrac{d\psi (t)}{dt}=g(t,0,\alpha )+\lambda , &\mbox{ $t\in (\overline {T},T]$,} \\[8pt] \psi (0)=\psi (T). \end{cases} \end{equation}
\begin{equation} \begin{cases} \dfrac {1}{\psi(t)} \dfrac{d\psi (t)}{dt}=f(t,0,\alpha )+\lambda , \quad & \mbox{ $t\in (0,\overline {T}]$,} \\[8pt] \dfrac {1}{\psi(t)}\dfrac{d\psi (t)}{dt}=g(t,0,\alpha )+\lambda , &\mbox{ $t\in (\overline {T},T]$,} \\[8pt] \psi (0)=\psi (T). \end{cases} \end{equation}
Integrating the first equation of (2.3) with respect to
 $t$
from
$t$
from
 $0$
to
$0$
to
 $\overline {T}$
and the second from
$\overline {T}$
and the second from
 $\overline {T}$
to
$\overline {T}$
to
 $T$
, we obtain
$T$
, we obtain
 \begin{equation*} \ln \psi (\overline {T})-\ln \psi (0)=\int ^{\overline {T}}_{0} (f(t,0,\alpha )+\lambda )\,{\rm d}t \end{equation*}
\begin{equation*} \ln \psi (\overline {T})-\ln \psi (0)=\int ^{\overline {T}}_{0} (f(t,0,\alpha )+\lambda )\,{\rm d}t \end{equation*}
and
 \begin{equation*} \ln \psi (T)-\ln \psi (\overline {T})=\int ^{T}_{\overline {T}} (g(t,0,\alpha )+\lambda )\,{\rm d}t, \end{equation*}
\begin{equation*} \ln \psi (T)-\ln \psi (\overline {T})=\int ^{T}_{\overline {T}} (g(t,0,\alpha )+\lambda )\,{\rm d}t, \end{equation*}
respectively. Adding the above two equations and using
 $\psi (0)=\psi (T)$
, we get
$\psi (0)=\psi (T)$
, we get
 \begin{equation} \lambda T+\int ^{\overline {T}}_{0}f(t,0,\alpha )\,{\rm d}t+\int ^{T}_{\overline {T}} g(t,0,\alpha )\,{\rm d}t=0. \end{equation}
\begin{equation} \lambda T+\int ^{\overline {T}}_{0}f(t,0,\alpha )\,{\rm d}t+\int ^{T}_{\overline {T}} g(t,0,\alpha )\,{\rm d}t=0. \end{equation}
By (H), let
 $\omega ^*$
be the unique root of
$\omega ^*$
be the unique root of
 $\digamma (T,\overline {T},\alpha )=0$
, where
$\digamma (T,\overline {T},\alpha )=0$
, where
 $\omega$
is one of
$\omega$
is one of
 $T$
,
$T$
,
 $\overline {T},$
and
$\overline {T},$
and
 $\alpha$
and correspondingly,
$\alpha$
and correspondingly,
 $\omega ^{*}$
can be one of
$\omega ^{*}$
can be one of
 $T^{*}$
,
$T^{*}$
,
 $\overline {T}^{*},$
and
$\overline {T}^{*},$
and
 $\alpha ^{*},$
respectively. Moreover, we have that
$\alpha ^{*},$
respectively. Moreover, we have that
 $\lambda$
satisfying (2.4) has the property,
$\lambda$
satisfying (2.4) has the property,
 \begin{equation*} \lambda \begin{cases} \gt 0 & \mbox{if $\omega \lt \omega ^*$,} \\ =0 & \mbox{if $\omega =\omega ^*$,} \\ \lt 0\quad & \mbox{if $\omega \gt \omega ^*$.} \end{cases} \end{equation*}
\begin{equation*} \lambda \begin{cases} \gt 0 & \mbox{if $\omega \lt \omega ^*$,} \\ =0 & \mbox{if $\omega =\omega ^*$,} \\ \lt 0\quad & \mbox{if $\omega \gt \omega ^*$.} \end{cases} \end{equation*}
Now, we are ready to give the main results of this paper.
We first present a result on the existence and attractivity of positive
 $T$
-periodic solutions to system (1.2), that is,
$T$
-periodic solutions to system (1.2), that is,
 \begin{equation} \begin{cases} \dfrac{{\rm d} x}{{\rm d} t}=xf(t,x,\alpha ), \quad &t\in (0,\overline {T}], \\[8pt] \dfrac{{\rm d} x}{{\rm d} t}=xg(t,x,\alpha ),&t\in (\overline {T},T], \\[8pt] x(0)=x(T). \end{cases} \end{equation}
\begin{equation} \begin{cases} \dfrac{{\rm d} x}{{\rm d} t}=xf(t,x,\alpha ), \quad &t\in (0,\overline {T}], \\[8pt] \dfrac{{\rm d} x}{{\rm d} t}=xg(t,x,\alpha ),&t\in (\overline {T},T], \\[8pt] x(0)=x(T). \end{cases} \end{equation}
Theorem 2.1.
Suppose that (H2) holds. If
 $\omega \gt \omega ^{*}$
, then system (1.2) has at least one positive
$\omega \gt \omega ^{*}$
, then system (1.2) has at least one positive
 $T$
-periodic solution. Furthermore, if (H3) holds, then system (1.2) has a unique globally attractive positive
$T$
-periodic solution. Furthermore, if (H3) holds, then system (1.2) has a unique globally attractive positive
 $T$
-periodic solution.
$T$
-periodic solution.
Proof.
Let
 $\psi (t)$
be a positive eigenfunction of (2.2) associated with the eigenvalue
$\psi (t)$
be a positive eigenfunction of (2.2) associated with the eigenvalue
 $\lambda$
satisfying (2.4). Denote
$\lambda$
satisfying (2.4). Denote
 $\underline {x}(t)=\epsilon \psi (t)$
. Since
$\underline {x}(t)=\epsilon \psi (t)$
. Since
 $\omega \gt \omega ^{*}$
, we have
$\omega \gt \omega ^{*}$
, we have
 $\lambda \lt 0$
. For small enough
$\lambda \lt 0$
. For small enough
 $\epsilon \gt 0$
, we obtain
$\epsilon \gt 0$
, we obtain
 \begin{equation*} \begin{cases} \phantom {=}\dfrac {{\rm d} \underline {x}}{{\rm d}t}-\underline {x}f(t,\underline {x},\alpha ) -\underline {x}f(t,0,\alpha )+\underline {x}f(t,0,\alpha ) \\[4pt] =({-}f(t,\epsilon \psi (t),\alpha )+f(t,0,\alpha )+\lambda )\epsilon \psi (t)\leq 0\quad & \mbox{for }t\in (0,\overline {T}], \\[4pt] \phantom {=}\dfrac {{\rm d} \underline {x}}{{\rm d}t}-\underline {x}g(t,\underline {x},\alpha ) -\underline {x}g(t,0,\alpha )+\underline {x}g(t,0,\alpha ) \\[4pt] =({-}g(t,\epsilon \psi (t),\alpha )+g(t,0,\alpha )+\lambda )\epsilon \psi (t)\leq 0\quad & \mbox{for }t\in (\overline {T},T], \\[4pt] \phantom {=}\psi (0)=\psi (T). \end{cases} \end{equation*}
\begin{equation*} \begin{cases} \phantom {=}\dfrac {{\rm d} \underline {x}}{{\rm d}t}-\underline {x}f(t,\underline {x},\alpha ) -\underline {x}f(t,0,\alpha )+\underline {x}f(t,0,\alpha ) \\[4pt] =({-}f(t,\epsilon \psi (t),\alpha )+f(t,0,\alpha )+\lambda )\epsilon \psi (t)\leq 0\quad & \mbox{for }t\in (0,\overline {T}], \\[4pt] \phantom {=}\dfrac {{\rm d} \underline {x}}{{\rm d}t}-\underline {x}g(t,\underline {x},\alpha ) -\underline {x}g(t,0,\alpha )+\underline {x}g(t,0,\alpha ) \\[4pt] =({-}g(t,\epsilon \psi (t),\alpha )+g(t,0,\alpha )+\lambda )\epsilon \psi (t)\leq 0\quad & \mbox{for }t\in (\overline {T},T], \\[4pt] \phantom {=}\psi (0)=\psi (T). \end{cases} \end{equation*}
Thus,
 $\epsilon \psi$
is a sub-solution of the periodic system (2.5). On the other hand, with the help of (H2), we can deduce that
$\epsilon \psi$
is a sub-solution of the periodic system (2.5). On the other hand, with the help of (H2), we can deduce that
 \begin{equation*} \begin{cases} \phantom {=} \dfrac {{\rm d} \bar {M}}{{\rm d} t}-\bar {M}f(t,\bar {M},\alpha ) \\[6pt] =-\bar {M}f(t,\bar {M},\alpha )\geq 0 \quad & \mbox{for }t\in (0,\overline {T}], \\[6pt] \phantom {=} \dfrac {{\rm d} \bar {M}}{{\rm d} t}-\bar {M}g(t,\bar {M},\alpha ) \\[6pt] =-\bar {M}g(t,\bar {M},\alpha )\geq 0\quad & \mbox{for }t\in (\overline {T},T] \end{cases} \end{equation*}
\begin{equation*} \begin{cases} \phantom {=} \dfrac {{\rm d} \bar {M}}{{\rm d} t}-\bar {M}f(t,\bar {M},\alpha ) \\[6pt] =-\bar {M}f(t,\bar {M},\alpha )\geq 0 \quad & \mbox{for }t\in (0,\overline {T}], \\[6pt] \phantom {=} \dfrac {{\rm d} \bar {M}}{{\rm d} t}-\bar {M}g(t,\bar {M},\alpha ) \\[6pt] =-\bar {M}g(t,\bar {M},\alpha )\geq 0\quad & \mbox{for }t\in (\overline {T},T] \end{cases} \end{equation*}
for
 $\bar {M}\geq \max \{M_{1},M_{2}\}$
. Therefore,
$\bar {M}\geq \max \{M_{1},M_{2}\}$
. Therefore,
 $(\bar {M},\epsilon \psi (t))$
forms a pair of upper and lower solutions for the periodic system (2.5). This implies that (2.5) has a positive
$(\bar {M},\epsilon \psi (t))$
forms a pair of upper and lower solutions for the periodic system (2.5). This implies that (2.5) has a positive
 $T$
-periodic solution
$T$
-periodic solution
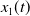 $x_{1}(t)$
initiated from the minimal point
$x_{1}(t)$
initiated from the minimal point
 $u_{1}$
.
$u_{1}$
.
Next, we prove that the periodic system (2.5) has a unique positive
 $T$
-periodic solution when (H3) also holds.
$T$
-periodic solution when (H3) also holds.
Suppose, for the sake of contradiction, that
 $x_2(t)$
is also a positive
$x_2(t)$
is also a positive
 $T$
-periodic solution to system (2.5) initiated from
$T$
-periodic solution to system (2.5) initiated from
 $u_{2}$
with
$u_{2}$
with
 $u_{1}\lt u_{2}$
. Since
$u_{1}\lt u_{2}$
. Since
 $x_{i}$
(
$x_{i}$
(
 $i=1$
,
$i=1$
,
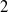 $2$
) is a positive solution to the following equation
$2$
) is a positive solution to the following equation
 \begin{equation*} \begin{cases} \dfrac {{\rm d} x}{{\rm d} t}=xf(t,x,\alpha ), \quad t\in (0,\overline {T}], \\[6pt] \dfrac {{\rm d} x}{{\rm d} t}=x g(t,x ,\alpha ), \quad t\in (\overline {T},T], \\[6pt] x (0)=(T), \end{cases} \end{equation*}
\begin{equation*} \begin{cases} \dfrac {{\rm d} x}{{\rm d} t}=xf(t,x,\alpha ), \quad t\in (0,\overline {T}], \\[6pt] \dfrac {{\rm d} x}{{\rm d} t}=x g(t,x ,\alpha ), \quad t\in (\overline {T},T], \\[6pt] x (0)=(T), \end{cases} \end{equation*}
we have
 \begin{equation} \begin{cases} \dfrac{1}{x_i}\dfrac{dx_i}{dt}=f(t,x_{i},\alpha ), \quad &t\in (0,\overline {T}], \\[6pt] \dfrac{1}{x_i}\dfrac{dx_i}{dt}=g(t,x_{i},\alpha ), & t\in (\overline {T},T]. \end{cases} \end{equation}
\begin{equation} \begin{cases} \dfrac{1}{x_i}\dfrac{dx_i}{dt}=f(t,x_{i},\alpha ), \quad &t\in (0,\overline {T}], \\[6pt] \dfrac{1}{x_i}\dfrac{dx_i}{dt}=g(t,x_{i},\alpha ), & t\in (\overline {T},T]. \end{cases} \end{equation}
Integrating the first equation of (2.6) in
 $t$
from
$t$
from
 $0$
to
$0$
to
 $\overline {T}$
and the second one from
$\overline {T}$
and the second one from
 $\overline {T}$
to
$\overline {T}$
to
 $T$
, and then summing the resultants, we obtain
$T$
, and then summing the resultants, we obtain
 \begin{equation*} \int ^{\overline {T}}_{0}f(t,x_{i},\alpha )\,{\rm d}t +\int ^{T}_{\overline {T}}g(t,x_{i},\alpha )\,{\rm d}t=0. \end{equation*}
\begin{equation*} \int ^{\overline {T}}_{0}f(t,x_{i},\alpha )\,{\rm d}t +\int ^{T}_{\overline {T}}g(t,x_{i},\alpha )\,{\rm d}t=0. \end{equation*}
According to (H3), this is impossible. Therefore, system (2.5) has a unique positive
 $T$
-periodic solution. Using a standard monotone iteration scheme and the instability of the trivial equilibrium point
$T$
-periodic solution. Using a standard monotone iteration scheme and the instability of the trivial equilibrium point
 $0$
of system (1.2), we see that the unique positive
$0$
of system (1.2), we see that the unique positive
 $T$
-periodic solution of system (1.2) is globally attractive. This completes the proof.
$T$
-periodic solution of system (1.2) is globally attractive. This completes the proof.
Next, we consider the extinction of system (1.2).
Theorem 2.2.
Suppose that (H1) holds. If
 $\omega \lt \omega ^{*}$
, then the trivial equilibrium point
$\omega \lt \omega ^{*}$
, then the trivial equilibrium point
 $0$
of system (1.2) is globally asymptotically stable.
$0$
of system (1.2) is globally asymptotically stable.
Proof.
Let
 $x(t)$
be a solution of (1.2). Define
$x(t)$
be a solution of (1.2). Define
 $\bar {x}=Me^{-\lambda t}\psi (t)$
, where
$\bar {x}=Me^{-\lambda t}\psi (t)$
, where
 $M$
is chosen large enough such that
$M$
is chosen large enough such that
 $x(0)\leq \bar {x}(0)$
and
$x(0)\leq \bar {x}(0)$
and
 $\psi$
is the one chosen in the proof of Theorem2.1. Then we have
$\psi$
is the one chosen in the proof of Theorem2.1. Then we have
 \begin{equation*} \begin{cases} \phantom {=}\dfrac {{\rm d} \bar {x}}{{\rm d} t}-\bar {x}f(t,\bar {x},\alpha )-\bar {x}f(t,0,\alpha )+\bar {x}f(t,0,\alpha ) \\[6pt] =({-}f(t,\bar {x},\alpha )+f(t,0,\alpha ))\bar {x}\geq 0, \quad & t\in (nT,nT+\overline {T}], \\[6pt] \phantom {=}\dfrac {{\rm d} \bar {x}}{{\rm d} t}-\bar {x}g(t,\bar {x},\alpha )-\bar {x}g(t,0,\alpha )+\bar {x}g(t,0,\alpha ) \\[6pt] =({-}g(t,\bar {x},\alpha )+g(t,0,\alpha ))\bar {x}\geq 0, & t\in (nT+\overline {T},(n+1)T], \\[6pt] \phantom {=}\bar {x}(0)\geq x(0), \end{cases} \end{equation*}
\begin{equation*} \begin{cases} \phantom {=}\dfrac {{\rm d} \bar {x}}{{\rm d} t}-\bar {x}f(t,\bar {x},\alpha )-\bar {x}f(t,0,\alpha )+\bar {x}f(t,0,\alpha ) \\[6pt] =({-}f(t,\bar {x},\alpha )+f(t,0,\alpha ))\bar {x}\geq 0, \quad & t\in (nT,nT+\overline {T}], \\[6pt] \phantom {=}\dfrac {{\rm d} \bar {x}}{{\rm d} t}-\bar {x}g(t,\bar {x},\alpha )-\bar {x}g(t,0,\alpha )+\bar {x}g(t,0,\alpha ) \\[6pt] =({-}g(t,\bar {x},\alpha )+g(t,0,\alpha ))\bar {x}\geq 0, & t\in (nT+\overline {T},(n+1)T], \\[6pt] \phantom {=}\bar {x}(0)\geq x(0), \end{cases} \end{equation*}
where we have used (H1). By the comparison principle, we can obtain that
 $x(t)\leq \bar {x}(t)$
for all
$x(t)\leq \bar {x}(t)$
for all
 $t\gt 0$
. Note that
$t\gt 0$
. Note that
 $\omega \lt \omega ^{*}$
implies
$\omega \lt \omega ^{*}$
implies
 $\lambda \gt 0$
. It follows that
$\lambda \gt 0$
. It follows that
 $\bar {x}(t)\to 0$
exponentially as
$\bar {x}(t)\to 0$
exponentially as
 $t\to +\infty$
. Therefore,
$t\to +\infty$
. Therefore,
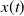 $x(t)$
must decay toward zero exponentially as well. This demonstrates that the trivial equilibrium point
$x(t)$
must decay toward zero exponentially as well. This demonstrates that the trivial equilibrium point
 $0$
is globally asymptotically stable.
$0$
is globally asymptotically stable.
The result below gives the asymptotical stability of
 $0$
when
$0$
when
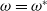 $\omega =\omega ^*$
under a little stronger condition.
$\omega =\omega ^*$
under a little stronger condition.
Theorem 2.3.
Suppose that (H1) holds and the inequalities there are strict. If
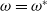 $\omega =\omega ^{*}$
, then the trivial equilibrium point
$\omega =\omega ^{*}$
, then the trivial equilibrium point
 $0$
of system (1.2) is globally asymptotically stable.
$0$
of system (1.2) is globally asymptotically stable.
Proof.
We first show that there is no positive
 $T$
-periodic solution to system (1.2). Assume, by contradiction, that there exists
$T$
-periodic solution to system (1.2). Assume, by contradiction, that there exists
 $\bar {x}(t)\gt 0$
for
$\bar {x}(t)\gt 0$
for
 $t\in [0,T]$
satisfying system (2.5), that is,
$t\in [0,T]$
satisfying system (2.5), that is,
 \begin{equation*} \begin{cases} \dfrac {{\rm d} \bar {x}}{{\rm d} t}=\bar {x}f(t,\bar {x},\alpha ), \quad &t\in (0,\overline {T}], \\[8pt] \dfrac {{\rm d} \bar {x}}{{\rm d} t}=\bar {x}g(t,\bar {x},\alpha ), &t\in (\overline {T},T], \\[8pt] \bar {x}(0)=\bar {x}(T). \end{cases} \end{equation*}
\begin{equation*} \begin{cases} \dfrac {{\rm d} \bar {x}}{{\rm d} t}=\bar {x}f(t,\bar {x},\alpha ), \quad &t\in (0,\overline {T}], \\[8pt] \dfrac {{\rm d} \bar {x}}{{\rm d} t}=\bar {x}g(t,\bar {x},\alpha ), &t\in (\overline {T},T], \\[8pt] \bar {x}(0)=\bar {x}(T). \end{cases} \end{equation*}
or, equivalently
 \begin{equation*} \begin{cases} \dfrac {1}{\bar{x}}\dfrac{d\bar{x}}{dt}=f(t,\bar {x},\alpha ),\quad &t\in (0,\overline {T}], \\[8pt] \dfrac {1}{\bar{x}}\dfrac{d\bar{x}}{dt}=g(t,\bar {x},\alpha ), & t\in (\overline {T},T]. \end{cases} \end{equation*}
\begin{equation*} \begin{cases} \dfrac {1}{\bar{x}}\dfrac{d\bar{x}}{dt}=f(t,\bar {x},\alpha ),\quad &t\in (0,\overline {T}], \\[8pt] \dfrac {1}{\bar{x}}\dfrac{d\bar{x}}{dt}=g(t,\bar {x},\alpha ), & t\in (\overline {T},T]. \end{cases} \end{equation*}
It follows that
 \begin{equation*} \int ^{\overline {T}}_{0}f(t,\bar {x},\alpha )\,{\rm d}t +\int ^{T}_{0}g(t,\bar {x},\alpha )\,{\rm d}t=0. \end{equation*}
\begin{equation*} \int ^{\overline {T}}_{0}f(t,\bar {x},\alpha )\,{\rm d}t +\int ^{T}_{0}g(t,\bar {x},\alpha )\,{\rm d}t=0. \end{equation*}
This combined with (2.4) implies that
 \begin{equation*} \lambda T=\int ^{\overline {T}}_{0}[f(t,\bar {x},\alpha ) -f(t,0,\alpha )]\,{\rm d}t+\int ^{T}_{0}[g(t,\bar {x},\alpha )-g(t,0,\alpha )]\,{\rm d}t. \end{equation*}
\begin{equation*} \lambda T=\int ^{\overline {T}}_{0}[f(t,\bar {x},\alpha ) -f(t,0,\alpha )]\,{\rm d}t+\int ^{T}_{0}[g(t,\bar {x},\alpha )-g(t,0,\alpha )]\,{\rm d}t. \end{equation*}
By the assumption that
 $f(t,0,\alpha )\gt f(t,x,\alpha )$
and
$f(t,0,\alpha )\gt f(t,x,\alpha )$
and
 $g(t,0,\alpha )\gt g(t,x,\alpha )$
for any
$g(t,0,\alpha )\gt g(t,x,\alpha )$
for any
 $x\gt 0$
, we obtain that
$x\gt 0$
, we obtain that
 $\lambda \lt 0$
. However, since
$\lambda \lt 0$
. However, since
 $\omega =\omega ^{*}$
, we have
$\omega =\omega ^{*}$
, we have
 $\lambda =0$
. This leads to a contradiction, and hence (1.2) has no positive
$\lambda =0$
. This leads to a contradiction, and hence (1.2) has no positive
 $T$
-periodic solution.
$T$
-periodic solution.
By a standard monotone iteration scheme and the nonexistence of positive
 $T$
-periodic solution of system (1.2), the trivial equilibrium point
$T$
-periodic solution of system (1.2), the trivial equilibrium point
 $0$
of system (1.2) is globally asymptotically stable. This completes the proof.
$0$
of system (1.2) is globally asymptotically stable. This completes the proof.
3 Two illustrating examples
In this section, we apply the results established in Section 2 to two switching systems from population dynamics.
3.1 A harvesting model
Due to overfishing, many fish resources are facing depletion worldwide, which has broken the balance of ecosystems. This has led to changes in the populations of other species and a weakening of overall ecosystem functions. As a response, many countries have implemented fishing bans to restore and protect marine biological resources, ensure ecosystem stability and diversity, and safeguard the livelihoods of fishermen as well as future food for humans.
However, a complete ban on all forms of fishing, though potentially beneficial for resource recovery, also presents several potential problems and negative impacts. Such a ban could lead to significant socioeconomic issues, particularly in communities that rely heavily on fishing as it is their primary economic activity. These communities may face severe livelihood losses. Additionally, fish are a critical source of protein for millions of people, especially in some poor countries. A total fishing ban could reduce their nutritional intake, potentially leading to malnutrition and associated health problems.
Therefore, developing reasonable fishing ban strategies is crucial for policymakers. The works [Reference Feng, Liu, Ruan and Yu6, Reference Liu, Feng, Ruan and Yu11, Reference Liu, Yu and Li13, Reference Xiao23] have explored how to determine appropriate fishing ban thresholds based on varying fishing intensities. Following the idea in [Reference Feng, Liu, Ruan and Yu6, Reference Liu, Feng, Ruan and Yu11], the main objective of this subsection is to understand the dynamics of the following problem,
 \begin{equation} \begin{cases} \dfrac {{\rm d}u}{{\rm d}t}=ru\left (1-\dfrac {u}{G}\right ), & {t\in (nT,nT+\overline {T}],} \\[8pt] \dfrac {{\rm d}u}{{\rm d}t}=ru\left (1-\dfrac {u}{G}\right ) -\dfrac {q(t)E(t)u}{cE(t)+lu}, \quad & {t\in (nT+\overline {T},(n+1)T],} \end{cases} \end{equation}
\begin{equation} \begin{cases} \dfrac {{\rm d}u}{{\rm d}t}=ru\left (1-\dfrac {u}{G}\right ), & {t\in (nT,nT+\overline {T}],} \\[8pt] \dfrac {{\rm d}u}{{\rm d}t}=ru\left (1-\dfrac {u}{G}\right ) -\dfrac {q(t)E(t)u}{cE(t)+lu}, \quad & {t\in (nT+\overline {T},(n+1)T],} \end{cases} \end{equation}
where
 $u(t)$
represents the population size of a single species at time
$u(t)$
represents the population size of a single species at time
 $t$
.
$t$
.
 $T$
is the period of the seasonal fluctuation environment where the single species lives;
$T$
is the period of the seasonal fluctuation environment where the single species lives;
 $\overline {T}$
measures the length of the closed season (non-harvesting season) and hence
$\overline {T}$
measures the length of the closed season (non-harvesting season) and hence
 $T-\overline {T}$
is the length of the open season (harvesting season); the parameter
$T-\overline {T}$
is the length of the open season (harvesting season); the parameter
 $r$
is the intrinsic growth rate of the species; and
$r$
is the intrinsic growth rate of the species; and
 $G$
is the carrying capacity of the environment. The term
$G$
is the carrying capacity of the environment. The term
 $\frac {q(t)E(t)u}{cE(t)+lu}$
is the harvesting rate, which is the Michaelis–Menten type functional form for the catch rate. Here
$\frac {q(t)E(t)u}{cE(t)+lu}$
is the harvesting rate, which is the Michaelis–Menten type functional form for the catch rate. Here
 $q(t)$
is the catch-ability coefficient at time
$q(t)$
is the catch-ability coefficient at time
 $t$
and
$t$
and
 $E(t)$
is the external effort devoted to harvesting at time
$E(t)$
is the external effort devoted to harvesting at time
 $t$
. We further assume that
$t$
. We further assume that
 $q(t)$
and
$q(t)$
and
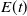 $E(t)$
are continuous and
$E(t)$
are continuous and
 $T$
-periodic in
$T$
-periodic in
 $t$
.
$t$
.
 $c$
and
$c$
and
 $l$
are two constants. All parameters are assumed to be non-negative.
$l$
are two constants. All parameters are assumed to be non-negative.
Corresponding to (1.2), here for (3.1), we have
 \begin{equation*} f(t,u,\alpha )=r\left (1-\frac {u}{G}\right ) \quad \mbox{and}\quad g(t,u,\alpha )=r\left (1-\frac {u}{G}\right )-\frac {q(t)E(t)}{cE(t)+lu}. \end{equation*}
\begin{equation*} f(t,u,\alpha )=r\left (1-\frac {u}{G}\right ) \quad \mbox{and}\quad g(t,u,\alpha )=r\left (1-\frac {u}{G}\right )-\frac {q(t)E(t)}{cE(t)+lu}. \end{equation*}
Then
 \begin{eqnarray*} \digamma (T,\overline {T},\alpha ) & =& \int _0^{\overline {T}}f(t,0,\alpha )\,{\rm d}t+\int _{\overline {T}}^T g(t,0,\alpha )\,{\rm d}t \notag \\[3pt] &=& \int _0^{\overline {T}}r\,{\rm d}t+\int _{\overline {T}}^T \left (r-\frac {q(t)}{c}\right )\,{\rm d}t \notag \\[3pt] & =& rT-\frac {1}{c}\int _{\overline {T}}^T q(t)\,{\rm d}t. \end{eqnarray*}
\begin{eqnarray*} \digamma (T,\overline {T},\alpha ) & =& \int _0^{\overline {T}}f(t,0,\alpha )\,{\rm d}t+\int _{\overline {T}}^T g(t,0,\alpha )\,{\rm d}t \notag \\[3pt] &=& \int _0^{\overline {T}}r\,{\rm d}t+\int _{\overline {T}}^T \left (r-\frac {q(t)}{c}\right )\,{\rm d}t \notag \\[3pt] & =& rT-\frac {1}{c}\int _{\overline {T}}^T q(t)\,{\rm d}t. \end{eqnarray*}
We first make some reasonable assumptions on the parameters. On the one hand, in the absence of fishing, that is,
 $T=\overline {T}$
, system (3.1) reduces to
$T=\overline {T}$
, system (3.1) reduces to
 \begin{equation*} \frac {{\rm d}u}{{\rm d}t}=ru\left (1-\frac {u}{G}\right ), \quad t\gt 0. \end{equation*}
\begin{equation*} \frac {{\rm d}u}{{\rm d}t}=ru\left (1-\frac {u}{G}\right ), \quad t\gt 0. \end{equation*}
It is necessary to assume that the population is persistent in the absence of fishing and hence
 $r\gt 0$
. On the other hand, when there is no fishing ban, that is,
$r\gt 0$
. On the other hand, when there is no fishing ban, that is,
 $\overline {T}=0$
, system (3.1) reduces to
$\overline {T}=0$
, system (3.1) reduces to
 \begin{equation*} \frac {{\rm d}u}{{\rm d}t}=ru\left (1-\frac {u}{G}\right ) -\frac {q(t)E(t)u}{cE(t)+lu}, \quad t\gt 0. \end{equation*}
\begin{equation*} \frac {{\rm d}u}{{\rm d}t}=ru\left (1-\frac {u}{G}\right ) -\frac {q(t)E(t)u}{cE(t)+lu}, \quad t\gt 0. \end{equation*}
Let
 $\inf _{t\in [\overline {T},T]}E(t)=E_{0}$
and
$\inf _{t\in [\overline {T},T]}E(t)=E_{0}$
and
 $\inf _{t\in [\overline {T},T]}q(t)=q_{0}$
. Then
$\inf _{t\in [\overline {T},T]}q(t)=q_{0}$
. Then
 \begin{equation*}\frac {{\rm d}u}{{\rm d}t}=ru\left (1-\frac {u}{G}\right )-\frac {q(t)E(t)u}{cE(t)+lu}\leq ru\left (1-\frac {u}{G}\right )-\frac {q_{0}E_{0}u}{cE_{0}+lu}. \end{equation*}
\begin{equation*}\frac {{\rm d}u}{{\rm d}t}=ru\left (1-\frac {u}{G}\right )-\frac {q(t)E(t)u}{cE(t)+lu}\leq ru\left (1-\frac {u}{G}\right )-\frac {q_{0}E_{0}u}{cE_{0}+lu}. \end{equation*}
Consider the system
 \begin{equation} \frac {{\rm d} \bar {u}}{{\rm d} t}=r\bar {u}\left (1-\frac {\bar {u}}{G}\right ) -\frac {q_{0}E_{0}\bar {u}}{cE_{0}+l\bar {u}}. \end{equation}
\begin{equation} \frac {{\rm d} \bar {u}}{{\rm d} t}=r\bar {u}\left (1-\frac {\bar {u}}{G}\right ) -\frac {q_{0}E_{0}\bar {u}}{cE_{0}+l\bar {u}}. \end{equation}
By comparison principle,
 $u(t)\le \bar {u}(t)$
, where
$u(t)\le \bar {u}(t)$
, where
 $u$
is the solution of system (3.1) and
$u$
is the solution of system (3.1) and
 $\bar {u}(t)$
is the solution of (3.2) with
$\bar {u}(t)$
is the solution of (3.2) with
 $u(0)=\bar {u}(0)$
. Clearly,
$u(0)=\bar {u}(0)$
. Clearly,
 $\bar {u}=0$
is the trivial equilibrium of (3.2) and its other positive equilibria are determined by the quadratic equation
$\bar {u}=0$
is the trivial equilibrium of (3.2) and its other positive equilibria are determined by the quadratic equation
 \begin{equation} \frac {rl}{G}\bar {u}^{2} +\left (\frac {rcE_{0}}{G}-rl\right )\bar {u}+(q_{0}-cr)E_{0}=0. \end{equation}
\begin{equation} \frac {rl}{G}\bar {u}^{2} +\left (\frac {rcE_{0}}{G}-rl\right )\bar {u}+(q_{0}-cr)E_{0}=0. \end{equation}
It is clear that equation (3.3) has no positive solution if
 \begin{equation} E_{0} \geq \frac {Gl}{c} \quad \mbox{and}\quad q_{0}\geq cr. \end{equation}
\begin{equation} E_{0} \geq \frac {Gl}{c} \quad \mbox{and}\quad q_{0}\geq cr. \end{equation}
Thus, if (3.4) holds, then
 $\bar {u}(t)\to 0$
and hence
$\bar {u}(t)\to 0$
and hence
 $u(t)\to 0$
as
$u(t)\to 0$
as
 $t\to +\infty$
. This means that the population will become extinct without fishing ban. Based on the above discussion, throughout the remainder of this subsection, for system (3.1), we always assume that (3.4) holds.
$t\to +\infty$
. This means that the population will become extinct without fishing ban. Based on the above discussion, throughout the remainder of this subsection, for system (3.1), we always assume that (3.4) holds.
Clearly,
 $\digamma (T,\overline {T},\alpha )$
is monotonically increasing in
$\digamma (T,\overline {T},\alpha )$
is monotonically increasing in
 $\overline {T}$
for fixed
$\overline {T}$
for fixed
 $T$
and
$T$
and
 $\alpha$
. Moreover,
$\alpha$
. Moreover,
 \begin{equation*} \digamma (T,0,\alpha )=rT-\frac {1}{c}\int _0^T q(t)\,{\rm d}t \le rT-\frac {q_0}{c}T\leq 0 \end{equation*}
\begin{equation*} \digamma (T,0,\alpha )=rT-\frac {1}{c}\int _0^T q(t)\,{\rm d}t \le rT-\frac {q_0}{c}T\leq 0 \end{equation*}
by (3.4) and
 \begin{equation*} \digamma (T,T,\alpha )=rT\gt 0 \end{equation*}
\begin{equation*} \digamma (T,T,\alpha )=rT\gt 0 \end{equation*}
Thus (H)(b) is satisfied. Let
 $\overline {T}^*$
be the unique non-negative root of
$\overline {T}^*$
be the unique non-negative root of
 $\digamma (T,\overline {T},\alpha )=0$
with fixed
$\digamma (T,\overline {T},\alpha )=0$
with fixed
 $T$
and
$T$
and
 $\alpha$
. Actually,
$\alpha$
. Actually,
 $\overline {T}^{*}=\frac {\bar {q}-cr}{\bar {q}}$
, where
$\overline {T}^{*}=\frac {\bar {q}-cr}{\bar {q}}$
, where
 $\bar {q}=\frac {\int ^{T}_{\overline {T}}q(t)\,\text{d}t}{T-\overline {T}}$
.
$\bar {q}=\frac {\int ^{T}_{\overline {T}}q(t)\,\text{d}t}{T-\overline {T}}$
.
Viewed from different perspectives, the fishing ban policy in high seas is usually a pre-established norm by countries, mainly aimed at protecting the breeding of marine fish during the breeding season. This means that the closing period
 $\overline {T}$
is fixed. Hence, from (H)(c), we have
$\overline {T}$
is fixed. Hence, from (H)(c), we have
 $r^{*}=\frac {(T-\overline {T})\bar {q}}{T}$
for fixed
$r^{*}=\frac {(T-\overline {T})\bar {q}}{T}$
for fixed
 $T$
and
$T$
and
 $\overline {T}$
. In this case, the parameter
$\overline {T}$
. In this case, the parameter
 $\alpha$
in system (1.2) is the parameter
$\alpha$
in system (1.2) is the parameter
 $r$
. However, in this subsection, we only considered the case of the threshold
$r$
. However, in this subsection, we only considered the case of the threshold
 $\overline {T}^{*}$
. The case of threshold
$\overline {T}^{*}$
. The case of threshold
 $r^{*}$
can be handled in a similar manner, and hence it is omitted here.
$r^{*}$
can be handled in a similar manner, and hence it is omitted here.
Now, we are ready to apply the results in Section 2 to system (3.1).
Theorem 3.1.
Suppose that
 $\overline {T}\gt \overline {T}^{*}$
. Then system (3.1) has at least one positive
$\overline {T}\gt \overline {T}^{*}$
. Then system (3.1) has at least one positive
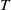 $T$
-periodic solution. Furthermore, if
$T$
-periodic solution. Furthermore, if
 $E_{0}\gt \frac {q^{0}lG}{rc^{2}}$
, then system (3.1) has a unique positive
$E_{0}\gt \frac {q^{0}lG}{rc^{2}}$
, then system (3.1) has a unique positive
 $T$
-periodic solution, which is globally asymptotically stable, where
$T$
-periodic solution, which is globally asymptotically stable, where
 $q^{0}\,:\!=\,\sup _{t\in [\overline {T}, T]} q(t)$
.
$q^{0}\,:\!=\,\sup _{t\in [\overline {T}, T]} q(t)$
.
Proof.
Clearly (H2) holds for
 $M_1=M_2=G$
and
$M_1=M_2=G$
and
 $f_u(t,u,\alpha )\lt 0$
for
$f_u(t,u,\alpha )\lt 0$
for
 $t\in (nT,nT+\overline {T}]$
and
$t\in (nT,nT+\overline {T}]$
and
 $u\gt 0$
. Moreover, when
$u\gt 0$
. Moreover, when
 $E_0\gt \frac {q^0lG}{rc^2}$
, for
$E_0\gt \frac {q^0lG}{rc^2}$
, for
 $t\in (nT+\overline {T}, (n+1)T]$
and
$t\in (nT+\overline {T}, (n+1)T]$
and
 $u\gt 0$
, we have
$u\gt 0$
, we have
 \begin{equation*} g_u(t,u,\alpha ) = -\frac {r}{G}+\frac {q(t)E(t)l}{(cE(t)+lu)^2} \le -\frac {r}{G}+\frac {q(t)E(t)l}{(cE(t))^2} \le -\frac {r}{G}+\frac {q^0l}{c^2E_0}\lt 0. \end{equation*}
\begin{equation*} g_u(t,u,\alpha ) = -\frac {r}{G}+\frac {q(t)E(t)l}{(cE(t)+lu)^2} \le -\frac {r}{G}+\frac {q(t)E(t)l}{(cE(t))^2} \le -\frac {r}{G}+\frac {q^0l}{c^2E_0}\lt 0. \end{equation*}
Thus (H3) holds if
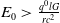 $E_0\gt \frac {q^0lG}{rc^2}$
. According to Theorem2.1, we are left to show that if
$E_0\gt \frac {q^0lG}{rc^2}$
. According to Theorem2.1, we are left to show that if
 $E_0\gt \frac {q^0lG}{rc^2}$
then the unique globally attractive positive
$E_0\gt \frac {q^0lG}{rc^2}$
then the unique globally attractive positive
 $T$
-periodic solution
$T$
-periodic solution
 $\bar {u}(t)$
is locally stable.
$\bar {u}(t)$
is locally stable.
Since
 $\bar {u}(t)$
satisfies
$\bar {u}(t)$
satisfies
 \begin{equation} \begin{cases} \dfrac {{\rm d}\bar {u}}{{\rm d}t}=r\bar {u}\left (1-\frac {\bar {u}}{G}\right ), & {t\in (0,\overline {T}],} \\[8pt] \dfrac {{\rm d}\bar {u}}{{\rm d}t}=r\bar {u}\left (1-\frac {\bar {u}}{G}\right ) -\dfrac {q(t)E(t)\bar {u}}{cE(t)+l\bar {u}}, \quad & t\in (\overline {T},T], \\[8pt] \bar {u}(0)=\bar {u}(T), \end{cases} \end{equation}
\begin{equation} \begin{cases} \dfrac {{\rm d}\bar {u}}{{\rm d}t}=r\bar {u}\left (1-\frac {\bar {u}}{G}\right ), & {t\in (0,\overline {T}],} \\[8pt] \dfrac {{\rm d}\bar {u}}{{\rm d}t}=r\bar {u}\left (1-\frac {\bar {u}}{G}\right ) -\dfrac {q(t)E(t)\bar {u}}{cE(t)+l\bar {u}}, \quad & t\in (\overline {T},T], \\[8pt] \bar {u}(0)=\bar {u}(T), \end{cases} \end{equation}
dividing the first two equations of (3.5) by
 $\bar {u}$
, then integrating the first resultant over
$\bar {u}$
, then integrating the first resultant over
 $(0,\overline {T}]$
and the latter over
$(0,\overline {T}]$
and the latter over
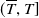 $(\overline {T},T]$
, and adding up the integrals, we obtain
$(\overline {T},T]$
, and adding up the integrals, we obtain
 \begin{equation*} \int ^{T}_{0}\frac {1}{\bar{u}}\frac{d\bar{u}}{dt} \,{\rm d}t = rT-r\int ^{T}_{0}\frac {\bar {u}}{G}\,{\rm d}t -\int ^{T}_{\overline {T}}\frac {q(t)E(t)}{cE(t)+l\bar {u}}\,{\rm d}t. \end{equation*}
\begin{equation*} \int ^{T}_{0}\frac {1}{\bar{u}}\frac{d\bar{u}}{dt} \,{\rm d}t = rT-r\int ^{T}_{0}\frac {\bar {u}}{G}\,{\rm d}t -\int ^{T}_{\overline {T}}\frac {q(t)E(t)}{cE(t)+l\bar {u}}\,{\rm d}t. \end{equation*}
Since
 $\bar {u}$
is
$\bar {u}$
is
 $T$
-periodic, we have
$T$
-periodic, we have
 $\int ^{T}_{0}\frac {1}{\bar{u}}\frac{d\bar{u}}{dt} \text{d}t=0$
and hence
$\int ^{T}_{0}\frac {1}{\bar{u}}\frac{d\bar{u}}{dt} \text{d}t=0$
and hence
 \begin{equation} rT-r\int ^{T}_{0}\frac {\bar {u}}{G}\,{\rm d}t -\int ^{T}_{\overline {T}}\frac {q(t)E(t)}{cE(t)+l\bar {u}}\,{\rm d}t=0. \end{equation}
\begin{equation} rT-r\int ^{T}_{0}\frac {\bar {u}}{G}\,{\rm d}t -\int ^{T}_{\overline {T}}\frac {q(t)E(t)}{cE(t)+l\bar {u}}\,{\rm d}t=0. \end{equation}
Let
 $v=u-\bar {u}$
. Then the linearised system of (3.1) about
$v=u-\bar {u}$
. Then the linearised system of (3.1) about
 $\bar {u}$
is
$\bar {u}$
is
 \begin{equation*} \begin{cases} \dfrac{{\rm d} v}{{\rm d} t}=rv\left (1-\dfrac {2\bar {u}}{G}\right ), & t\in (0,\overline {T}], \\[8pt] \dfrac{{\rm d} v}{{\rm d} t}=rv\left (1-\dfrac {2\bar {u}}{G}\right ) -\dfrac {cE^{2}(t)q(t) v}{(cE(t)+l \bar {u})^{2}}, \quad & t\in (\overline {T},T]. \end{cases} \end{equation*}
\begin{equation*} \begin{cases} \dfrac{{\rm d} v}{{\rm d} t}=rv\left (1-\dfrac {2\bar {u}}{G}\right ), & t\in (0,\overline {T}], \\[8pt] \dfrac{{\rm d} v}{{\rm d} t}=rv\left (1-\dfrac {2\bar {u}}{G}\right ) -\dfrac {cE^{2}(t)q(t) v}{(cE(t)+l \bar {u})^{2}}, \quad & t\in (\overline {T},T]. \end{cases} \end{equation*}
Let
 $v=e^{-\tau t}\varphi (t)$
with
$v=e^{-\tau t}\varphi (t)$
with
 $\varphi (0)=\varphi (T)$
. This leads to the following eigenvalue problem,
$\varphi (0)=\varphi (T)$
. This leads to the following eigenvalue problem,
 \begin{equation*} \begin{cases} \dfrac{{\rm d} \varphi }{{\rm d} t}=r\left (1-\dfrac {2\bar {u}}{G}\right )\varphi +\tau \varphi , & t\in (0,\overline {T}], \\[8pt] \dfrac{{\rm d} \varphi }{{\rm d} t}=r\left (1-\dfrac {2\bar {u}}{G}\right )\varphi -\dfrac {cE^{2}(t)q(t) \varphi }{(cE(t)+l \bar {u})^{2}}+\tau \varphi ,\quad & t\in (\overline {T},T], \\[8pt] \varphi (0)=\varphi (T). \end{cases} \end{equation*}
\begin{equation*} \begin{cases} \dfrac{{\rm d} \varphi }{{\rm d} t}=r\left (1-\dfrac {2\bar {u}}{G}\right )\varphi +\tau \varphi , & t\in (0,\overline {T}], \\[8pt] \dfrac{{\rm d} \varphi }{{\rm d} t}=r\left (1-\dfrac {2\bar {u}}{G}\right )\varphi -\dfrac {cE^{2}(t)q(t) \varphi }{(cE(t)+l \bar {u})^{2}}+\tau \varphi ,\quad & t\in (\overline {T},T], \\[8pt] \varphi (0)=\varphi (T). \end{cases} \end{equation*}
Similar to obtain (3.6), we can get
 \begin{equation} \tau T-\int ^{T}_{\overline {T}}\frac {cE^{2}(t)q(t)}{(cE(t)+l \bar {u})^{2}}\,{\rm d}t+rT-2\frac {r}{G}\int ^{T}_{0}\bar {u} \,{\rm d}t=0. \end{equation}
\begin{equation} \tau T-\int ^{T}_{\overline {T}}\frac {cE^{2}(t)q(t)}{(cE(t)+l \bar {u})^{2}}\,{\rm d}t+rT-2\frac {r}{G}\int ^{T}_{0}\bar {u} \,{\rm d}t=0. \end{equation}
It follows from (3.6) and (3.7) that
 \begin{eqnarray*} \tau T&=& \int ^{T}_{\overline {T}}\frac {cE^{2}(t)q(t)}{(cE(t)+l \bar {u})^{2}}\,{\rm d}t +rT-2\int ^{T}_{\overline {T}}\frac {q(t)E(t)}{cE(t)+l\bar {u}}\,{\rm d}t, \notag \\[4pt] &= & rT-\int ^{T}_{\overline {T}}\frac {cE^{2}(t)q(t)+2E(t)q(t)l\bar {u}}{(cE(t)+l \bar {u})^{2}}\,{\rm d}t, \notag \\[4pt] &\geq & rT-\int ^{T}_{\overline {T}}\frac {q(t)}{c}\,{\rm d}t \notag \\[4pt] &=&\frac {\bar {q}}{c}(\overline {T}-\overline {T}^{*}). \end{eqnarray*}
\begin{eqnarray*} \tau T&=& \int ^{T}_{\overline {T}}\frac {cE^{2}(t)q(t)}{(cE(t)+l \bar {u})^{2}}\,{\rm d}t +rT-2\int ^{T}_{\overline {T}}\frac {q(t)E(t)}{cE(t)+l\bar {u}}\,{\rm d}t, \notag \\[4pt] &= & rT-\int ^{T}_{\overline {T}}\frac {cE^{2}(t)q(t)+2E(t)q(t)l\bar {u}}{(cE(t)+l \bar {u})^{2}}\,{\rm d}t, \notag \\[4pt] &\geq & rT-\int ^{T}_{\overline {T}}\frac {q(t)}{c}\,{\rm d}t \notag \\[4pt] &=&\frac {\bar {q}}{c}(\overline {T}-\overline {T}^{*}). \end{eqnarray*}
Thus,
 $\tau \gt 0$
due to
$\tau \gt 0$
due to
 $\overline {T}\gt \overline {T}^{*}$
, which implies that
$\overline {T}\gt \overline {T}^{*}$
, which implies that
 $\bar {u}$
is locally stable. This completes the proof.
$\bar {u}$
is locally stable. This completes the proof.
Note that, for
 $u\gt 0$
,
$u\gt 0$
,
 \begin{equation*} f(t,0,\alpha )=r\gt r\left (1-\frac {u}{G}\right )=f(t,u,\alpha ) \end{equation*}
\begin{equation*} f(t,0,\alpha )=r\gt r\left (1-\frac {u}{G}\right )=f(t,u,\alpha ) \end{equation*}
and
 \begin{eqnarray*} g(t,0,\alpha )-g(t,u,\alpha )& = & u\left (\frac {r}{G} -\frac {q(t)l}{c(cE(t)+lu)}\right ) \notag \\[4pt] & \ge & u\left (\frac {r}{G}-\frac {q(t)l}{c^2E(t)}\right ) \notag \\[4pt] & \ge & u\left (\frac {r}{G}-\frac {q^0l}{c^2E_0}\right ). \end{eqnarray*}
\begin{eqnarray*} g(t,0,\alpha )-g(t,u,\alpha )& = & u\left (\frac {r}{G} -\frac {q(t)l}{c(cE(t)+lu)}\right ) \notag \\[4pt] & \ge & u\left (\frac {r}{G}-\frac {q(t)l}{c^2E(t)}\right ) \notag \\[4pt] & \ge & u\left (\frac {r}{G}-\frac {q^0l}{c^2E_0}\right ). \end{eqnarray*}
These combined with Theorem2.2 and Theorem2.3 produce the following result on the extinction of system (3.1).
Theorem 3.2.
The trivial equilibrium point
 $0$
of system (3.1) is globally asymptotically stable if either
$0$
of system (3.1) is globally asymptotically stable if either
 \begin{equation*} \overline {T}\lt \overline {T}^* \quad \mbox{and} \quad E_0\ge \frac {q^0lG}{rc^2} \end{equation*}
\begin{equation*} \overline {T}\lt \overline {T}^* \quad \mbox{and} \quad E_0\ge \frac {q^0lG}{rc^2} \end{equation*}
or
 \begin{equation*} \overline {T}=\overline {T}^* \quad \mbox{and} \quad E_0\gt \frac {q^0lG}{rc^2}. \end{equation*}
\begin{equation*} \overline {T}=\overline {T}^* \quad \mbox{and} \quad E_0\gt \frac {q^0lG}{rc^2}. \end{equation*}
3.2 A switching mosquito population suppression model
Mosquito-borne diseases are caused by bacteria, viruses or parasites transmitted to people by mosquitoes. Typical mosquito-borne diseases include Zika, West Nile, Chikungunya, dengue, yellow fever, and malaria. These diseases have become a serious threat to public health worldwide, which cause over 725,000 deaths and nearly 700 million infections each year [18]. Due to the lack of vaccines and effective clinical treatments for most of the above-mentioned mosquito-borne diseases, and the additional risks associated with inadequate preventive measures, mosquito vector control has become the most important strategy [Reference Daily3, Reference Godói, Lemos, de Araújo, Bonoto, Godman and Guerra Júnior7, Reference Schwartz, Halloran, Durbin and Longini19]. The traditional measure is the application of chemical insecticides. This not only brings some environment problems and public health issues but also has limited effects on depressing the mosquito population as mosquitoes can quickly develop insecticide resistance [Reference Kyle and Harris10, Reference Ooi, Goh and Gubler14]. Therefore, scientists have started to develop environment-friendly insect control measures, including Sterile insect technique (SIT) and Incompatible insect technology (IIT).
SIT is an environment-friendly insect control technology that aimed to reduce the population of target pests by releasing radiation-sterilised males into the field to mate with wild females so that these wild females cannot produce viable offsprings. In practice, SIT has been successfully applied in the control of several important pests in agriculture and animal husbandry [Reference Dyck, Hendrichs and Robinson5].
IIT is an approach that releases male mosquitoes infected with the maternally inherited endosymbiotic bacteria Wolbachia into the field, resulting in cytoplasmic incompatibility so that the matings with wild females that are not infected with the same Wolbachia strain become sterile – thereby suppressing the mosquito population [Reference Turelli and Hoffmann21, Reference Xi, Khoo and Dobson22]. Wolbachia is an endosymbiotic bacterium that is parasitised exclusively in invertebrates and can be transmitted through eggs [Reference Pfarr, Foster and Slatko17]. It is estimated that about
 $65\%$
insect species and
$65\%$
insect species and
 $28\%$
mosquito species naturally carry Wolbachia [Reference Hilgenboecker, Hammerstein and Schlattmann8, Reference Pattamaporn, Baisley, Visut and O’Neill16]. IIT is very promising since it is not only environment friendly but also very efficient with strong specificity and low cost. In 2005, a group led by Xi successfully transplanted Wolbachia carried by fruit flies into the germ cells of Aedes aegypti and made it spread stably in Aedes aegypti [Reference Xi, Khoo and Dobson22]. This made the mass release of millions of factory-reared incompatible adult Aedes aegypti technically feasible.
$28\%$
mosquito species naturally carry Wolbachia [Reference Hilgenboecker, Hammerstein and Schlattmann8, Reference Pattamaporn, Baisley, Visut and O’Neill16]. IIT is very promising since it is not only environment friendly but also very efficient with strong specificity and low cost. In 2005, a group led by Xi successfully transplanted Wolbachia carried by fruit flies into the germ cells of Aedes aegypti and made it spread stably in Aedes aegypti [Reference Xi, Khoo and Dobson22]. This made the mass release of millions of factory-reared incompatible adult Aedes aegypti technically feasible.
The basic principles of mosquito population suppression based on SIT and IIT are the same, both involving the release of sterile male mosquitoes in the target area to suppress the target mosquito population. In a recent work [Reference Yu and Li24], Yu and Li formulated and analysed the following interactive model of wild and sterile mosquitoes,
 \begin{equation} \frac{{\rm d} w}{{\rm d} t}=\left [\frac {aw}{w+g(t)}-(\mu +\xi (w+g(t)))\right ]w. \end{equation}
\begin{equation} \frac{{\rm d} w}{{\rm d} t}=\left [\frac {aw}{w+g(t)}-(\mu +\xi (w+g(t)))\right ]w. \end{equation}
Here
 $w$
is the number of wild male mosquitoes at time
$w$
is the number of wild male mosquitoes at time
 $t$
,
$t$
,
 $a$
represents the birth rate of wild mosquitoes,
$a$
represents the birth rate of wild mosquitoes,
 $\mu$
denotes the density-independent death rate and
$\mu$
denotes the density-independent death rate and
 $\xi$
is the density-dependent death coefficient. In this context,
$\xi$
is the density-dependent death coefficient. In this context,
 $g$
is treated as a given function rather than an independent variable governed by a dynamic equation. They assumed that a fixed number
$g$
is treated as a given function rather than an independent variable governed by a dynamic equation. They assumed that a fixed number
 $c$
of mosquitoes is released at regular intervals of length
$c$
of mosquitoes is released at regular intervals of length
 $T$
. Additionally, their research highlights the importance of a crucial parameter – the sexual lifespan, denoted by
$T$
. Additionally, their research highlights the importance of a crucial parameter – the sexual lifespan, denoted by
 $\overline {T}$
– which plays a significant role in mosquito suppression strategies. It is assumed that after the sexually active period, mosquitoes are no longer vigorous, and their competitive impact on wild mosquitoes becomes negligible, though alive. Under the assumption that
$\overline {T}$
– which plays a significant role in mosquito suppression strategies. It is assumed that after the sexually active period, mosquitoes are no longer vigorous, and their competitive impact on wild mosquitoes becomes negligible, though alive. Under the assumption that
 $g(t)$
becomes a piecewise constant
$g(t)$
becomes a piecewise constant
 $T$
-periodic function defined by
$T$
-periodic function defined by
 \begin{equation} \begin{aligned} g(t)=\left \{ \begin{array}{c@{\quad}l} c,\, &{t\in (nT,nT+\overline {T}]},\\[5pt] 0,\, &{t\in (nT+\overline {T},(n+1)T]}, \end{array} \right . \end{aligned} \end{equation}
\begin{equation} \begin{aligned} g(t)=\left \{ \begin{array}{c@{\quad}l} c,\, &{t\in (nT,nT+\overline {T}]},\\[5pt] 0,\, &{t\in (nT+\overline {T},(n+1)T]}, \end{array} \right . \end{aligned} \end{equation}
for
 $n=0$
,
$n=0$
,
 $1$
,
$1$
,
 $2$
,
$2$
,
 $\ldots$
, system (3.8) becomes
$\ldots$
, system (3.8) becomes
 \begin{equation*}\begin{cases} \dfrac{{\rm d} w}{{\rm d} t}=w\left (\dfrac {aw}{w+c}-(\mu +\xi (w+c))\right ),\quad &t\in (nT,nT+\overline {T}], \\[8pt] \dfrac{{\rm d} w}{{\rm d} t}=w\left (a-\mu -\xi w\right ), &t\in (nT+\overline {T},(n+1)T]. \end{cases} \end{equation*}
\begin{equation*}\begin{cases} \dfrac{{\rm d} w}{{\rm d} t}=w\left (\dfrac {aw}{w+c}-(\mu +\xi (w+c))\right ),\quad &t\in (nT,nT+\overline {T}], \\[8pt] \dfrac{{\rm d} w}{{\rm d} t}=w\left (a-\mu -\xi w\right ), &t\in (nT+\overline {T},(n+1)T]. \end{cases} \end{equation*}
One assumption that cannot be overlooked in [Reference Yu and Li24] is that mosquitoes released during the period
 $(nT,nT+\overline {T}]$
will not die. In this subsection, we propose a more reasonable assumption: mosquitoes do die during their sexual lifespan. As a result, the constant
$(nT,nT+\overline {T}]$
will not die. In this subsection, we propose a more reasonable assumption: mosquitoes do die during their sexual lifespan. As a result, the constant
 $c$
in (3.9) should be a monotonically decreasing function of
$c$
in (3.9) should be a monotonically decreasing function of
 $t$
with period
$t$
with period
 $T$
. This leads to the following model on mosquito suppression,
$T$
. This leads to the following model on mosquito suppression,
 \begin{equation} \begin{cases} \dfrac{{\rm d} w}{{\rm d} t}=w\left (\frac {aw}{w+c(t)}-(\mu +\xi (w+c(t)))\right ), \quad &t\in (nT,nT+\overline {T}], \\[8pt] \dfrac{{\rm d} w}{{\rm d} t}=w\left (a-\mu -\xi w\right ), & t\in (nT+\overline {T},(n+1)T]. \end{cases} \end{equation}
\begin{equation} \begin{cases} \dfrac{{\rm d} w}{{\rm d} t}=w\left (\frac {aw}{w+c(t)}-(\mu +\xi (w+c(t)))\right ), \quad &t\in (nT,nT+\overline {T}], \\[8pt] \dfrac{{\rm d} w}{{\rm d} t}=w\left (a-\mu -\xi w\right ), & t\in (nT+\overline {T},(n+1)T]. \end{cases} \end{equation}
Similarly as in Section 3.1, on the one hand, we assume persistence of system (3.10) in the absence of sterile male mosquitoes, which leads to the condition that
 $a\gt \mu$
. On the other hand, we assume that extinction occurs when
$a\gt \mu$
. On the other hand, we assume that extinction occurs when
 $T=\overline {T}$
(meaning the release frequency equals the mosquito’s sexual lifespan). Under this assumption, system (3.10) becomes
$T=\overline {T}$
(meaning the release frequency equals the mosquito’s sexual lifespan). Under this assumption, system (3.10) becomes
 \begin{equation*} \frac{{\rm d} w}{{\rm d} t}=w\left (\frac {aw}{w+c(t)}-(\mu +\xi (w+c(t)))\right ), \quad t\gt 0. \end{equation*}
\begin{equation*} \frac{{\rm d} w}{{\rm d} t}=w\left (\frac {aw}{w+c(t)}-(\mu +\xi (w+c(t)))\right ), \quad t\gt 0. \end{equation*}
Since
 $\inf _{t\in [0,\overline {T}]}c(t)=c(\overline {T})$
, we have
$\inf _{t\in [0,\overline {T}]}c(t)=c(\overline {T})$
, we have
 \begin{equation*} \frac{{\rm d} w}{{\rm d} t}=w\left (\frac {aw}{w+c(t)}-(\mu +\xi (w+c(t)))\right )\leq w\left (\frac {aw}{w+c(\overline {T})}-(\mu +\xi (w+c(\overline {T})))\right ). \end{equation*}
\begin{equation*} \frac{{\rm d} w}{{\rm d} t}=w\left (\frac {aw}{w+c(t)}-(\mu +\xi (w+c(t)))\right )\leq w\left (\frac {aw}{w+c(\overline {T})}-(\mu +\xi (w+c(\overline {T})))\right ). \end{equation*}
Now, consider the following auxiliary equation,
 \begin{equation} \frac{{\rm d} w}{{\rm d} t}=w\left (\frac {aw}{w+c(\overline {T})}-(\mu +\xi (w+c(\overline {T})))\right ),\quad t\gt 0. \end{equation}
\begin{equation} \frac{{\rm d} w}{{\rm d} t}=w\left (\frac {aw}{w+c(\overline {T})}-(\mu +\xi (w+c(\overline {T})))\right ),\quad t\gt 0. \end{equation}
When
 $c(\overline {T})\gt g^{*}\,:\!=\,\frac {(a-\mu )^{2}}{4\xi a}$
,
$c(\overline {T})\gt g^{*}\,:\!=\,\frac {(a-\mu )^{2}}{4\xi a}$
,
 $0$
is the only equilibrium of (3.11) and it is globally asymptotically stable. By the comparison principle, under this assumption, the mosquito population will be extinct. As a result, we assume that
$0$
is the only equilibrium of (3.11) and it is globally asymptotically stable. By the comparison principle, under this assumption, the mosquito population will be extinct. As a result, we assume that
 \begin{equation*}a\gt \mu \quad \mbox{and} \quad c(\overline {T})\gt g^*. \end{equation*}
\begin{equation*}a\gt \mu \quad \mbox{and} \quad c(\overline {T})\gt g^*. \end{equation*}
In terms of (1.2), for (3.10), we have
 \begin{equation*} f(t,w,\alpha )=\frac {aw}{w+c(t)}-(\mu +\xi (w+c(t))), \quad g(t,w,\alpha )=a-\mu -\xi w. \end{equation*}
\begin{equation*} f(t,w,\alpha )=\frac {aw}{w+c(t)}-(\mu +\xi (w+c(t))), \quad g(t,w,\alpha )=a-\mu -\xi w. \end{equation*}
Thus,
 \begin{eqnarray*} \digamma (T,\overline {T},\alpha ) & = & \int _0^{\overline {T}}f(t,0,\alpha )\,{\rm d}t +\int _{\overline {T}}^T g(t,0,\alpha )\,{\rm d}t \notag \\[4pt] & = & \int _0^{\overline {T}}[-(\mu +\xi c(t))]\,{\rm d}t+\int _{\overline {T}}^T (a-\mu )\,{\rm d}t \notag \\[4pt] &=& -\mu T+a(T-\overline {T})-\xi \int _0^{\overline {T}}c(t)\,{\rm d}t. \end{eqnarray*}
\begin{eqnarray*} \digamma (T,\overline {T},\alpha ) & = & \int _0^{\overline {T}}f(t,0,\alpha )\,{\rm d}t +\int _{\overline {T}}^T g(t,0,\alpha )\,{\rm d}t \notag \\[4pt] & = & \int _0^{\overline {T}}[-(\mu +\xi c(t))]\,{\rm d}t+\int _{\overline {T}}^T (a-\mu )\,{\rm d}t \notag \\[4pt] &=& -\mu T+a(T-\overline {T})-\xi \int _0^{\overline {T}}c(t)\,{\rm d}t. \end{eqnarray*}
Clearly, for fixed
 $\overline {T}\gt 0$
and
$\overline {T}\gt 0$
and
 $\alpha \gt 0$
,
$\alpha \gt 0$
,
 $\digamma (T,\overline {T},\alpha )$
is strictly monotonically increasing in
$\digamma (T,\overline {T},\alpha )$
is strictly monotonically increasing in
 $T$
,
$T$
,
 \begin{equation*} \digamma (\overline {T},\overline {T},\alpha )=-\mu \overline {T}-\xi \int _0^{\overline {T}}c(t)\,{\rm d}t\lt 0, \end{equation*}
\begin{equation*} \digamma (\overline {T},\overline {T},\alpha )=-\mu \overline {T}-\xi \int _0^{\overline {T}}c(t)\,{\rm d}t\lt 0, \end{equation*}
and
 $\lim \limits _{T\to +\infty }\digamma (T,\overline {T},\alpha )=+\infty$
as
$\lim \limits _{T\to +\infty }\digamma (T,\overline {T},\alpha )=+\infty$
as
 $a\gt \mu$
. Thus, (H)(b) holds. Let
$a\gt \mu$
. Thus, (H)(b) holds. Let
 $T^*$
be the unique positive root of
$T^*$
be the unique positive root of
 $\digamma (T,\overline {T},\alpha )=0$
, that is,
$\digamma (T,\overline {T},\alpha )=0$
, that is,
 \begin{equation*} T^*=\frac {a\overline {T}+\xi \int _0^{\overline {T}}c(t)\,{\rm d}t}{a-\mu }. \end{equation*}
\begin{equation*} T^*=\frac {a\overline {T}+\xi \int _0^{\overline {T}}c(t)\,{\rm d}t}{a-\mu }. \end{equation*}
One can easily check that
 \begin{equation*} f(t,w,\alpha )\le 0 \quad \textrm{for}\ w\ge M_1\,:\!=\,\frac {a}{\xi c(\overline {T})} \end{equation*}
\begin{equation*} f(t,w,\alpha )\le 0 \quad \textrm{for}\ w\ge M_1\,:\!=\,\frac {a}{\xi c(\overline {T})} \end{equation*}
and
 \begin{equation*} g(t,w,\alpha )\le 0 \quad \textrm{for}\ w\ge M_2\,:\!=\,\frac {a}{\xi }, \end{equation*}
\begin{equation*} g(t,w,\alpha )\le 0 \quad \textrm{for}\ w\ge M_2\,:\!=\,\frac {a}{\xi }, \end{equation*}
that is, (H2) holds. Moreover, for
 $w\gt 0$
,
$w\gt 0$
,
 \begin{equation*} f_w(t,w,\alpha )=\frac {ac(t)}{(w+c(t))^2}-\xi \lt \frac {a}{c(t)}-\xi \le \frac {a}{c(\overline {T})}-\xi \end{equation*}
\begin{equation*} f_w(t,w,\alpha )=\frac {ac(t)}{(w+c(t))^2}-\xi \lt \frac {a}{c(t)}-\xi \le \frac {a}{c(\overline {T})}-\xi \end{equation*}
and
 \begin{equation*} g_w(t,w,\alpha )=-\xi \lt 0. \end{equation*}
\begin{equation*} g_w(t,w,\alpha )=-\xi \lt 0. \end{equation*}
It follows that (H3) holds if
 $c(\overline {T})\ge \frac {a}{\xi }$
. According to Theorem2.1 and arguing similarly as in the proof of Theorem3.1 for local stability, we have the following result on the existence and stability of
$c(\overline {T})\ge \frac {a}{\xi }$
. According to Theorem2.1 and arguing similarly as in the proof of Theorem3.1 for local stability, we have the following result on the existence and stability of
 $T$
-periodic solutions to (3.10).
$T$
-periodic solutions to (3.10).
Theorem 3.3.
Suppose
 $T\gt T^{*}$
. Then system 3.10) has at least one positive
$T\gt T^{*}$
. Then system 3.10) has at least one positive
 $T$
-periodic solution. Furthermore, if
$T$
-periodic solution. Furthermore, if
 $c(\overline {T})\geq \frac {a}{\xi }$
, then it has a unique positive
$c(\overline {T})\geq \frac {a}{\xi }$
, then it has a unique positive
 $T$
-periodic solution, which is globally asymptotically stable.
$T$
-periodic solution, which is globally asymptotically stable.
Now, for
 $w\gt 0$
, we observe that
$w\gt 0$
, we observe that
 \begin{eqnarray*} f(t,0,\alpha )-f(t,w,\alpha )& = & w\left ({-}\frac {a}{w+c(t)}+\xi \right ) \\[2pt] & \gt & w\left ({-}\frac {a}{c(t)}+\xi \right ) \\[2pt] & \ge & w\left ({-}\frac {a}{c(\overline {T})}+\xi \right ) \\[2pt] & \gt & 0 \quad \left(\textrm{if}\ c(\overline {T})\ge \frac {a}{\xi }\right) \end{eqnarray*}
\begin{eqnarray*} f(t,0,\alpha )-f(t,w,\alpha )& = & w\left ({-}\frac {a}{w+c(t)}+\xi \right ) \\[2pt] & \gt & w\left ({-}\frac {a}{c(t)}+\xi \right ) \\[2pt] & \ge & w\left ({-}\frac {a}{c(\overline {T})}+\xi \right ) \\[2pt] & \gt & 0 \quad \left(\textrm{if}\ c(\overline {T})\ge \frac {a}{\xi }\right) \end{eqnarray*}
and
 \begin{equation*} g(t,0,\alpha )=a-\mu \gt a-\mu -\xi w=g(t,w,\alpha ). \end{equation*}
\begin{equation*} g(t,0,\alpha )=a-\mu \gt a-\mu -\xi w=g(t,w,\alpha ). \end{equation*}
Therefore, if
 $c(\overline {T})\ge \frac {a}{\xi }$
then (H1) holds and the inequalities there are strict ones. Applying Theorems2.2 and 2.3 to (3.10) produces the following result on extinction.
$c(\overline {T})\ge \frac {a}{\xi }$
then (H1) holds and the inequalities there are strict ones. Applying Theorems2.2 and 2.3 to (3.10) produces the following result on extinction.
Theorem 3.4.
Assume that
 $T\le T^{*}$
and
$T\le T^{*}$
and
 $c(\overline {T})\geq \frac {a}{\xi }$
. Then the trivial equilibrium
$c(\overline {T})\geq \frac {a}{\xi }$
. Then the trivial equilibrium
 $0$
of system (3.10) is globally asymptotically stable.
$0$
of system (3.10) is globally asymptotically stable.
Finally, we provide some numerical simulations to support the theoretical results obtained for system (3.10). We take
 \begin{equation*} a=10, \quad \mu =0.3, \quad \xi =0.2, \quad \overline {T}=2.5. \end{equation*}
\begin{equation*} a=10, \quad \mu =0.3, \quad \xi =0.2, \quad \overline {T}=2.5. \end{equation*}
With
 $c(t)=32.5-2t$
for
$c(t)=32.5-2t$
for
 $t\in [0,\overline {T}]$
, we have
$t\in [0,\overline {T}]$
, we have
 $c(\overline {T})=27.5\gt \frac {a}{\xi }=11.7612$
. Corresponding to this release strategy, the threshold of release waiting period is
$c(\overline {T})=27.5\gt \frac {a}{\xi }=11.7612$
. Corresponding to this release strategy, the threshold of release waiting period is
 $T^{*}=\frac {a\overline {T}+\xi \int ^{\overline {T}}_{0}c(t)\,{\rm d}t} {(a-\mu )}=400/97\approx 4.124$
. If the release waiting period
$T^{*}=\frac {a\overline {T}+\xi \int ^{\overline {T}}_{0}c(t)\,{\rm d}t} {(a-\mu )}=400/97\approx 4.124$
. If the release waiting period
 $T=4.5 \gt T^*$
, it follows from Theorem3.3 that system (3.10) has a unique globally asymptotically stable positive
$T=4.5 \gt T^*$
, it follows from Theorem3.3 that system (3.10) has a unique globally asymptotically stable positive
 $T$
-periodic solution as shown in Figure 1. If
$T$
-periodic solution as shown in Figure 1. If
 $T\le T^*$
, then Theorem3.4 tells us that the origin is globally asymptotically stable for (3.10). Figure 2 is for the case where
$T\le T^*$
, then Theorem3.4 tells us that the origin is globally asymptotically stable for (3.10). Figure 2 is for the case where
 $T=T^*$
whereas Figure 3 shows the case
$T=T^*$
whereas Figure 3 shows the case
 $T=4.024\lt T^*$
. The initial values for the figures in Figure 2 are chosen from the intervals
$T=4.024\lt T^*$
. The initial values for the figures in Figure 2 are chosen from the intervals
 $[10,30]$
,
$[10,30]$
,
 $[5,10)$
,
$[5,10)$
,
 $[1,4]$
, and
$[1,4]$
, and
 $(0,1]$
, respectively. Though the rate approaching
$(0,1]$
, respectively. Though the rate approaching
 $0$
is somewhat slow, they do indicate the convergence to
$0$
is somewhat slow, they do indicate the convergence to
 $0$
.
$0$
.

Figure 1 When
 $T\gt T^*$
and
$T\gt T^*$
and
 $c(\overline {T})\ge \frac {a}{\xi }$
, (3.10) has a globally asymptotically stable positive
$c(\overline {T})\ge \frac {a}{\xi }$
, (3.10) has a globally asymptotically stable positive
 $T$
-periodic solution.
$T$
-periodic solution.

Figure 2 When
 $T=T^*$
and
$T=T^*$
and
 $c(\overline {T})\ge \frac {a}{\xi }$
, the origin is globally asymptotically stable for (3.10). The initial values for the four figures are chosen form the four intervals,
$c(\overline {T})\ge \frac {a}{\xi }$
, the origin is globally asymptotically stable for (3.10). The initial values for the four figures are chosen form the four intervals,
 $[10,30]$
,
$[10,30]$
,
 $[5,10)$
,
$[5,10)$
,
 $(1,4]$
, and
$(1,4]$
, and
 $(0,1]$
. From the four figures, it can be seen that the origin is stable, although its approach to the origin is somewhat slow.
$(0,1]$
. From the four figures, it can be seen that the origin is stable, although its approach to the origin is somewhat slow.

Figure 3 When
 $T\lt T^*$
and
$T\lt T^*$
and
 $c(\overline {T})\ge \frac {a}{\xi }$
, the origin is globally asymptotically stable for (3.10).
$c(\overline {T})\ge \frac {a}{\xi }$
, the origin is globally asymptotically stable for (3.10).
4 Conclusion and discussion
The evolution of many biological processes are affected by seasonal factors. In this paper, we proposed a general periodic switching system to describe such phenomena. Under some reasonable assumptions, we established a threshold dynamics, which is completely characterised by a threshold value
 $\omega ^*$
. Roughly speaking, when
$\omega ^*$
. Roughly speaking, when
 $\omega \gt \omega ^*$
, there is a positive periodic solution, which is globally attractive under further conditions; when
$\omega \gt \omega ^*$
, there is a positive periodic solution, which is globally attractive under further conditions; when
 $\omega \leq \omega ^*$
, the origin is globally asymptotically stable.
$\omega \leq \omega ^*$
, the origin is globally asymptotically stable.
We also applied the theoretical results to two situations. One is for a population model with harvesting and the other is for mosquito control. For the harvesting model, we obtained a globally asymptotically stable positive periodic solution when
 $\bar {T}\gt \bar {T}^*$
. This implies that for resource sustainability, the harvesting should be banned at least for a certain time during a cycle. While for the mosquito suppression model, for a better control result, the waiting period for releasing Wolbachia-infected male mosquitoes should not be too large.
$\bar {T}\gt \bar {T}^*$
. This implies that for resource sustainability, the harvesting should be banned at least for a certain time during a cycle. While for the mosquito suppression model, for a better control result, the waiting period for releasing Wolbachia-infected male mosquitoes should not be too large.
We mention that the obtained theoretical results for the general system (1.2) is applicable to other situations. To end this paper, we briefly introduce an SIS model without conducting the detailed analysis. The classical SIS model is described by the system of ordinary differential equations,
 \begin{equation*} \begin{cases} \dfrac{{\rm d}S}{{\rm d}t}=-\beta f(S,I)+\gamma I, \\[8pt] \dfrac{{\rm d}I}{{\rm d}t}=\beta f(S,I)-\gamma I, \end{cases} \end{equation*}
\begin{equation*} \begin{cases} \dfrac{{\rm d}S}{{\rm d}t}=-\beta f(S,I)+\gamma I, \\[8pt] \dfrac{{\rm d}I}{{\rm d}t}=\beta f(S,I)-\gamma I, \end{cases} \end{equation*}
where
 $S(t)$
and
$S(t)$
and
 $I(t)$
are the numbers of susceptible and infectious individuals at time
$I(t)$
are the numbers of susceptible and infectious individuals at time
 $t$
, respectively. Here
$t$
, respectively. Here
 $f(S,I)$
is the incidence,
$f(S,I)$
is the incidence,
 $\beta$
is the infection rate, and
$\beta$
is the infection rate, and
 $\gamma$
is the recovery rate. Seasonal succession, the start of the school season and holiday breaks periodically alter behaviour patterns of people whereas environmental conditions and medical resources affect the infection and recovery rates [Reference Tang, Xiao, Zhang and Zhu20]. With these factors in consideration, we introduce the following modified periodic SIS model,
$\gamma$
is the recovery rate. Seasonal succession, the start of the school season and holiday breaks periodically alter behaviour patterns of people whereas environmental conditions and medical resources affect the infection and recovery rates [Reference Tang, Xiao, Zhang and Zhu20]. With these factors in consideration, we introduce the following modified periodic SIS model,
 \begin{equation} \begin{cases} \dfrac{{\rm d}S}{{\rm d}t}=-\beta (t)f(S,I)+\gamma I, \\[8pt] \dfrac{{\rm d}I}{{\rm d}t}= \beta (t)f(S,I)-\gamma I. \end{cases} \end{equation}
\begin{equation} \begin{cases} \dfrac{{\rm d}S}{{\rm d}t}=-\beta (t)f(S,I)+\gamma I, \\[8pt] \dfrac{{\rm d}I}{{\rm d}t}= \beta (t)f(S,I)-\gamma I. \end{cases} \end{equation}
It is easy to see that
 $\frac{{\rm d} (S+I)}{{\rm d}t}=0$
. This means that the total population size
$\frac{{\rm d} (S+I)}{{\rm d}t}=0$
. This means that the total population size
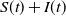 $S(t)+I(t)$
is a constant and is denoted by
$S(t)+I(t)$
is a constant and is denoted by
 $N$
. Then (4.1) is reduced to
$N$
. Then (4.1) is reduced to
 \begin{equation} \frac{{\rm d}I}{{\rm d}t}= \beta (t)f(N-I,I)-\gamma I. \end{equation}
\begin{equation} \frac{{\rm d}I}{{\rm d}t}= \beta (t)f(N-I,I)-\gamma I. \end{equation}
Now, we assume that the infection rate fluctuate periodically due to seasonal changes (such as temperature variations) as well as the effects of the school season and holiday breaks. Consequently, the infection rate exhibits cyclical alternations, that is, we assume
 \begin{equation*} \beta (t)= \begin{cases} \beta _{1}(t), &{t\in (nT,nT+\overline {T}]}, \\[5pt] \beta _{2}(t), \quad &{t\in (nT+\overline {T},(n+1)T]} \end{cases} \end{equation*}
\begin{equation*} \beta (t)= \begin{cases} \beta _{1}(t), &{t\in (nT,nT+\overline {T}]}, \\[5pt] \beta _{2}(t), \quad &{t\in (nT+\overline {T},(n+1)T]} \end{cases} \end{equation*}
for
 $n=0$
,
$n=0$
,
 $1$
,
$1$
,
 $2$
,
$2$
,
 $\ldots$
. Here
$\ldots$
. Here
 $T$
is the period of the disease transmissions. Due to variations in weather temperature and timing factors such as the school season and holidays, the parameter
$T$
is the period of the disease transmissions. Due to variations in weather temperature and timing factors such as the school season and holidays, the parameter
 $\overline {T}\in (0,T)$
is used to describe the transition points between different seasons. Similar to the analysis in the preceding sections, under certain assumptions, we can derive the threshold value of the recovery rate
$\overline {T}\in (0,T)$
is used to describe the transition points between different seasons. Similar to the analysis in the preceding sections, under certain assumptions, we can derive the threshold value of the recovery rate
 $\gamma$
based on assumption (H).
$\gamma$
based on assumption (H).
Financial support
This research was supported partially by the National Natural Science Foundation of China [12471458, 12171110, 12331017, 12071095], the Natural Sciences and Engineering Research Council of Canada [RGPIN-2024-05593], and the Basic and Applied Basic Research Foundation of Guangdong Province [2023A1515011110].
Competing interests
Authors state no conflict of interest.
Data availability statement
Our manuscript has no associated data.















 Open access
Open access